
Abstract
Infectious diseases pose major socioeconomic and health-related threats to millions of people across the globe. Strategies to combat infectious diseases derive from our understanding of the complex interactions between the host and specific bacterial, viral, and fungal pathogens. Lipid rafts are membrane microdomains that play important role in life cycle of microbes. Interaction of microbial pathogens with host membrane rafts influences not only their initial colonization but also their spread and the induction of inflammation. Therefore, intervention strategies aimed at modulating the assembly of membrane rafts and/or regulating raft-directed signaling pathways are attractive approaches for the. management of infectious diseases. The current review discusses the latest advances in terms of techniques used to study the role of membrane microdomains in various pathological conditions and provides updated information regarding the role of membrane rafts during bacterial, viral and fungal infections.
Similar content being viewed by others


Introduction
The cell, the basic unit of life, is separated from its surroundings by a selectively permeable cell membrane. Receptors and channels within cell membranes mediate interactions between environmental factors and cells. Early concepts of cell membranes envisioned a thin film of lipoidal material coating water-soluble proteins and surrounding the living cell. This model, which allowed for selective membrane permeability, was superseded by the fluid mosaic model of Singer and Nicholson (Singer and Nicolson 1972). According to the fluid mosaic model, cell membranes are two-dimensional liquid organelles with random distribution of phospholipids and proteins in no specific organization pattern. Since introduction of this concept, there have been revolutionary changes regarding the concept of the plasma membrane and this has been an area of intense research. For example, the existence of detergent-resistant sphingolipid-rich domains was demonstrated by Yu et al. (1973) and asymmetric distribution of phospholipids in the plasma membrane of erythrocytes were demonstrated by van Meer and coworkers (van Meer et al. 1980, 1987). Further, Simons and Ikonen (1997) (Simons and Ikonen 1997) proposed a glycosphingolipid/cholesterol raft model based on the formation of lateral lipid assemblies in an unsaturated glycerophospholipid environment. Since then, several reports have shown that the plasma membrane is actually more mosaic than fluid (Pike 2003; Engelman 2005; Vieira et al. 2010), and that the lipids are not randomly distributed. This has lead to the current model in which the plasma membrane is patchy, with segregated portions that are distinct in structure and function and may vary in thickness and composition (Pike 2003; Garcia-Marcos et al. 2006; Pike 2009). The formation of rafts is thought to be mediated by attractive forces between sphingolipids with saturated hydrocarbon chains and cholesterol. This model is based on the fact that formation of lipid rafts cannot be based only on lipid-protein interactions as it also depends on lipid-lipid associations, which includes phase separation coupled with lateral associations. These associations involve formation of hydrogen bonds, electrostatic forces, van der Waals forces, hydrophobic, and hydrophilic interactions (Pike 2009; Riethmuller et al. 2006; Nicolson 2014). In a seminal talk given at the Keystone Symposium on Lipid Rafts and Cell Function (2006) in Steamboat Springs, CO, lipid rafts were defined as small (10–200 nm), heterogeneous, highly dynamic, and sterol- and sphingolipid-rich domains that compartmentalize cellular processes (Pike 2006). At this Keynote Symposium, it was also suggested that small rafts can sometimes be stabilized to form larger platforms through protein-protein and protein-lipid interactions.
Lipid rafts are enriched in sphingolipids like sphingomyelin and cholesterol and have a preponderance of saturated acyl chains (Mishra and Joshi 2007; Lingwood and Simons 2010). Membrane proteins are attached via glycosylphosphatidylinositol (GPI) anchors to the lipid raft entities. The signaling proteins which reside or translocate to the rafts include Ras, nitric oxide synthase (NOS), phosphatidylinositol 3-kinase (PI3K), tyrosine kinases including Fyn and Src, insulin, and epidermal growth factors to name a few. These membrane domains/rafts exert their actions by separating or concentrating specific membrane proteins and lipids within microdomains, which were earlier thought to serve as platforms in the trans-Golgi network for apical membrane sorting and as foci for the recruitment and concentration of signaling molecules at the plasma membrane (Alonso and Millan 2001). The receptors and stimuli that induce the formation of rafts include cluster of differentiation (CD)5, CD20, CD40, CD95, bacterial and viral infections, as well as exposure to UV light, γ-irradiation, and chemical agents (Bollinger et al. 2005). Numerous proteins involved in cell signaling have been identified in caveolae and lipid rafts (planar), suggesting that these structures function as signal transduction moieties. Depletion of membrane cholesterol through the use of cholesterol-binding drugs or by blocking cellular cholesterol biosynthesis disrupts the formation and function of both lipid rafts and caveolae, indicating that these membrane domains are involved in a wide range of biological processes. The role of lipid rafts in bringing together specific set of proteins in response to stimulus has been extensively documented in the literature (Simons and Toomre 2000; Lafont and van der Goot 2005). Further, lipid rafts can also include or exclude proteins depending on their modifications. For example, palmitoylation increases the affinity of proteins for rafts, but not the ability of proteins to associate with rafts (Melkonian et al. 1999). Another example is monomeric transmembrane proteins which only reside in rafts for time periods until they are crosslinked or oligomerized (Harder et al. 1998). The distribution of lipids between the lipid bilayer has also been shown to play an important role in membrane function (Simons and Toomre 2000).
Caveolins and flotillins constitute a group of proteins that are enriched within lipid rafts (Hansen and Nichols 2009; Bauer and Pelkmans 2006; Yao et al. 2009). Caveolin-1, a palmitoylated membrane protein, is an essential component of a major subclass of rafts known as caveolae (flask-shaped invaginations of the plasma membrane), which causes rafts to polymerize (Rothberg et al. 1992; Anderson 1998). In contrast, planar rafts have flotillins as essential components (Giri et al. 2007; Zhao et al. 2011). Three caveolin isoforms transcribed from different genes have been identified so far which include caveolin-1, caveolin-2, and caveolin-3. The role of caveolin-1 and caveolin-3 in the formation of caveolae is well documented; however, the role of caveolin-2 is unclear (Sowa et al. 2008; Drab et al. 2001; Razani et al. 2001). Caveolin-1 mediates activation of insulin receptor-β and regulation of the angiotensin/TGF-β pathways suggesting a role in diabetes (Yamamoto et al. 1998) and pulmonary and cardiac fibrosis, (Wang et al. 2006; Cohen et al. 2003), respectively. On the other hand, caveolin-3 defects are associated with muscular dystrophy (autosomal dominant limb girdle muscular dystrophy, idiopathic hyperCKemia, rippling muscle disease, and distal myopathy) (Minetti et al. 1998; Betz et al. 2001; Tateyama et al. 2002). The flotillin family includes flotillin-1 and flotillin-2, which are important for signaling, endocytosis, and interactions of rafts with the cytoskeleton. Functional roles of flotillins have also been documented in polarization and chemotaxis of neutrophils in response to chemo attractants in vivo (Ludwig et al. 2010; Rossy et al. 2009). Both types of raft entities are also enriched in cholesterol and glycosphingolipids (Simons and Ikonen 1997; Korade and Kenworthy 2008; Wang 2014). The outer leaflet consists of sphingolipids, phosphatidylcholine, and sphingomyelin, whereas the inner leaflet consists of phosphatidylinositol, phosphatidylethanolamine, and phosphatidylserine with cholesterol being found in both leaflets (Mishra and Joshi 2007). Enrichment of phospholipids with saturated fatty acids allows close packing of lipids within rafts, and as a result lipid rafts are more complex and less fluid than the surrounding membrane.
Meanwhile, the formation of rafts rich in ceramide following exposure to external stimuli/stress has also received significant attention (Hannun and Obeid 2002) (Tables 1 and 2). Ceramide can be generated by de novo synthesis or hydrolysis of sphingomyelin by sphingomyelinases (Gault et al. 2010; Silva et al. 2009). Ceramide is composed of sphingosine and a fatty acid chain, and is found in higher concentrations within the cell membrane. An increasing body of literature has shown ceramide (sphingolipid), as an important molecule that regulates a diverse array of cellular processes including differentiation, immune responses, apoptosis, growth arrest, and senescence (Korade and Kenworthy 2008; Caliceti et al. 2012; Arana et al. 2010; DiNitto et al. 2003; Castro et al. 2009; Megha and London 2004; Yu et al. 2005). The role of sphingomyelinase and ceramide has been implicated in pathological conditions including pulmonary failure and erythrocyte apoptosis caused by platelet activating factor (Goggel et al. 2004; Lang et al. 2005). Furthermore, ceramide is also involved in the fusion of endosomes with lysosomes during the internalization of microbial pathogens into mammalian cells (Bollinger et al. 2005; Anes et al. 2003; Gulbins et al. 2004; Miller et al. 2012; Hartlova et al. 2010; Heung et al. 2006). Conversion of sphingomyelin into ceramide plays an important role in modulation of the membrane structure, which results in membrane vesiculation, fission/fusion and vesicular trafficking and contributes to cellular signaling (Devaux et al. 2008). Interestingly, rafts on one leaflet of the membrane can induce localized changes in the other leaflet of the bilayer, and they can potentially serve as the link between signals from outside the cell to those produced inside the cell (van Meer et al. 2008). In addition, ceramide has been shown to form organized, large channels traversing the mitochondrial outer membrane, which leads to the egress of proteins from the intermembrane space. Ceramide may affect the permeability of the mitochondrial outer membrane and the release of cytochrome C (Siskind et al. 2002). Thus, in the process of apoptosis, the breakdown of plasma membrane sphingomyelin to ceramide results in disorganization of rafts and may regulate apoptotic body formation. High ceramide levels in the lipid rafts increases the size of these platforms due to fusion, and reorganized ceramide-rich rafts retain/restrict proteins differently than other rafts (Spiegel and Milstien 2003). Ceramide-activated proteins act as secondary messengers and they are directly involved in activation of protein phosphatases PP1 and PP2, isoforms of protein kinase C, cathepsin D, and phospholipase A2, other signaling pathway components, although the precise mechanism of activation is unknown yet. Thus, formation of ceramide may serve different functions at distinct locations in the cell, and therefore, the effects of ceramide rafts are still under investigation (Silva et al. 2009; Zheng et al. 2006).
Techniques used to study the formation and functions of lipid rafts
Ordered and tightly packed acyl chains in lipid raft domains render them the ability to withstand disruption by non-ionic detergents, whereas the low protein/lipid ratio gives them much lower density than other solubilized membrane proteins. Utilizing this property, lipid rafts were originally isolated by sucrose density gradient ultracentrifugation from cellular extracts prepared in 1% Triton-X (Mishra and Joshi 2007; Ostrom and Liu 2007). Several other detergents have subsequently been used to isolate rafts. An alternative approach to study the role of lipid rafts or the association of certain components with rafts is treating them with cholesterol-sequestering agents or chelators like methyl-β-cyclodextrin to disrupt/deplete lipid raft constituents. Moreover, exogenous unsaturated fatty acids, gangliosides or cholesterol can also be used to destabilize raft assembly (Ostrom and Liu 2007; Brown 2006). The drawback of using a cholesterol-sequestering agent or chelator is the possible induction of structural or metabolic changes. Therefore, these may not be appropriate options to determine raft-associated cellular changes. Alternatively, genetic approaches such as small interfering RNA (siRNA) or gene knockout animals are widely used to disrupt planar rafts or caveolae (Pelkmans et al. 2004; Maguy et al. 2006). These studies address the physiological importance of membrane microdomains and help to determine their importance in signal transduction and bacterial/viral entry into cells (Zhong et al. 2008).
Recent use of more advanced and non-disruptive techniques have significantly advanced our understanding about these highly dynamic and fragile rafts. These include fluorescence correlation spectroscopy (FCS), single-molecule tracking microscopy (Kusumi et al. 2014; Parton and del Pozo 2013; Gambin et al. 2014), the use of a 2-dimethylamino-6-lauroylnaphthalene (laurdan) labeling probe for studying lateral organization of membranes using conventional confocal microscopy (Dodes Traian et al. 2012), and fluorescence resonance energy transfer (FRET) for determining the formation and influence of lipid rafts (Silvius and Nabi 2006). The sensitivity of the FCS technique for the determination of phase separation using free standing artificial lipid bilayers was demonstrated in earlier reports (Korlach et al. 1999; Bacia et al. 2004). This technique utilizes excitation and detection as in scanning microscopy to determine temporal fluorescence fluctuations due to the diffusion of individual fluorescent molecules. The autocorrelation curve generated from these fluctuations is used to determine the mobility (diffusion coefficients) of particles and can be used to monitor a single molecule to hundreds of molecules (Schutz et al. 2000; Ha 2001a, b). Further advancement in fluorescence microscopy techniques came with single molecule imaging (SMI) or single molecule tracking (SMT). This technique provides the ability to monitor collisions, interactions, binding, dissociation, organization, concentration, and disengagement of molecules in living cells with some limitations (Kusumi et al. 2014; Kusumi et al. 2005a; 2005b). Therefore, SMT, which is performed using high resolution photon detector(s) to track only one molecule or to simultaneously image thousands of molecules, has significantly advanced understanding of the three-tiered architecture of plasma membrane (Hinner and Johnsson 2010). In conjugation with FRET, SMT can also be used to detect activation of single molecules. The combination of these two techniques was first used to assess the activation of the G protein Ras (fluorescently labeled), which functions downstream of receptor-type tyrosine kinases, by detecting its binding the fluorescent GTP (energy acceptor) (Murakoshi et al. 2004). Moreover, a fluorescent membrane probe, Laurdan, can be used to determine the extent of water penetration into the lipid bilayer enabling assessment of lipid packing and membrane fluidity. The photophysical properties of laurdan determine its spectral sensitivity in polar solvents and leads to a shift of its emission spectra in loosely packed membranes with higher water penetrability (Dodes Traian et al. 2012; Kwiatek et al. 2013). The studies conducted using these techniques not only support the existence of rafts in live cells, but electron paramagnetic resonance studies and single molecule tracking methods have been used to determine the half-life of rafts and to assess their interactions with various proteins, respectively (Swamy et al. 2006). As a result, it is now clear that the half-life of rafts is in the range of 102 ns, which is not only faster than the rate of many enzyme reactions, but also less than the half-life of interactions between several proteins known to be involved in signal transduction (Swamy et al. 2006). Thus, there was imminent need for improving techniques for isolation of membrane rafts and for exploring the functional changes involved in signaling. In this regard, a recent study by Doughty et al. used secondary ion mass spectrometry (Nano SIMS) technology to determine the subcellular localization of hopanoid lipids in bacteria using stable-isotope labeling (Doughty et al. 2014). The SIMS technique, however, uses primary ions like Cs+ or O− to bombard the surface of sample resulting in generation of secondary ions that are then measured by mass spectrometry. Images are formed from the collection of mass spectra data from several spots on the surface. This technique can also be utilized to determine isotope ratios with high precision and used to visualize labeled sample regions (Altelaar et al. 2006; Grignon 2007). Furthermore, advances in microscopy techniques utilizing fluorescent labels or genetically-encoded probes and the mini-singlet–oxygen generator (miniSOG) to visualize the subcellular localization of proteins and lipids by electron microscopy moved the field forward significantly (Huang et al. 2009a). Additionally, new super-resolution microscopy techniques such as 3D-structured illumination microscopy (SIM), photoactivated localization microscopy (PALM), stochastic optical reconstruction (STORM), and stimulated emission depletion microscopy (STED) allow the investigation of lipid rafts at a level that was previously unattainable (Huang et al. 2009a). Among these, SIM is an ultra-resolution light microscopy technique that utilizes patterned illumination from a coherent light source to observe structures below the resolution limit of light microscopy by generating difference/beat frequencies termed Moiré fringes. This technique can be used with any conventional fluorophore and cell preparation (Kusumi et al. 2005a, b, 2014). On the other hand, PALM method is based on serial photoactivation or photoswitching and subsequent bleaching of several randomly scattered fluorescent proteins. This method provides a significant edge over the limitations of electron microscopy and conventional fluorescent microscopy to quantify protein aggregates or view cluster sizes that lie below diffraction limits. Utilizing total internal reflection fluorescence geometry (TIRF) in PALM further improves the detection range of fluorescent molecules. In order to investigate lipid rafts, another important technique which can be utilized is STORM imaging. This method involves several imaging cycles where a fraction of fluorophores are optically resolved in the field of view during each cycle, resulting in a highly accurate determination of individual active fluorophores (Rust et al. 2006). Yet another important technique in lipid rafts research is the STED technique, which uses non-linear fluorophore responses to improve the resolution and acquire images below the diffraction limit (Kusumi et al. 2014; Eggeling et al. 2013; Sahl and Moerner 2013). Both PALM and STORM techniques, on the contrary utilize mathematical modeling to reconstruct the sub-diffraction limit from multiple sets of diffraction-limited images (Rust et al. 2006; Sengupta et al. 2011; Hess et al. 2006). To summarize, the utilization of novel tools has helped generate deeper insight into the functional aspects of lipid rafts, which will help direct future studies and therapeutic intervention strategies targeting lipid rafts and lipid raft-associated proteins.
Lipid rafts and bacterial infections
Lipid rafts have been shown to play an important role during bacterial infections. Evidence of the ruffling of membrane lipids and proteins during pathogen engulfment and requirement for cholesterol during pathogen invasion substantiates this claim. Further, the requirement of raft-associated receptors on epithelial cells, including CD55 on the apical membrane and CD44 on the basolateral membrane, has been reported for bacterial entry (Selvarangan et al. 2000; Peiffer et al. 1998; Stuart et al. 2002a; Oliferenko et al. 1999). Several strains of bacteria invade phagocytes and non-phagocytic cells through the endocytic pathway in order to avoid the host defense mechanism (Simons and Gruenberg 2000; Steinberg and Grinstein 2008; Gruenberg and van der Goot 2006). Pathogens entering the host cells through the endosome-lysosome pathway involving clathrin-coated pits have the ability to avoid fusion or acidification following fusion with lysosomes to prevent degradation. Recent evidence suggests that these endocytic entry mechanisms are directed by the structure of lipid rafts, which are internalized via clathrin-dependent pathways. After endocytosis lipid rafts can form caveosomes and fuse with the Golgi complex or endoplasmic reticulum, or alternatively, form intracellular vesicles and remain within the organelle. The entry of pathogens via lipid rafts promotes intracellular survival and dissemination within the host. Raft-mediated entry of Escherichia coli (E. coli), Shigella flexneri, Salmonella thyphymurium, Mycobacterium spp., Chlamydia spp., Ehrlichia chaffeensis, Anaplasma phagocytophilum, Campylobacter jejuni (Lafont and van der Goot 2005; Manes et al. 2003a), Listeria monocytogenes (Seveau et al. 2004), Mycoplasma fermantas (Yavlovich et al. 2006), and Sphingomonas spp. (Ammendolia et al. 2004) have been studied extensively to find out the role of lipid rafts in the pathogenesis of the bacterial infections. In this regard, role of molecular motors kinesin and cytoplasmic dynein has also been investigated in terms of trafficking of pathogens inside cells towards lysosomal degradation. Molecular motors are important in the transport of organelle cargoes along microtubules in the cell. This transport driven by molecular motors, either unidirectional or bidirectional is regulated through their post-translational modifications, interactions with effector molecules or autoinhibition. Movement of early phagosomes along microtubules is bidirectional manner, and of late phagosomes is unidirectional which promotes its fusion with lysosomes (Grover et al. 2016). In a recent study, Rai et al. demonstrated that the mobility of early and late phagosomes correlates with the appearance of lipid rafts and clustering of dynein motors in phagosome membrane. The geometrical reorganization allows dyneins within cluster generate cooperative force on a single microtubule and results in rapid directed transport of phagosome which probably promotes phagosome and lysosome fusion leading to pathogen degradation. Disruption of clustering of small lipid microdomains by several pathogens can helps in survival of pathogen inside the host by inhibition of phagosome motion and phagosome-lysosome fusion (Rai et al. 2016). Interestingly, lysosome function can also be regulated by cholesterol. Further, Deng and coworkers demonstrated that cholesterol depletion can also disrupt lysosome membrane permeability (Deng et al. 2009). In this line of thought, mycobacteria was shown to exploit lipid rafts to enhance their adaptive fitness and protection from intracellular degradation by lysosomal pathways. Mycobacteria induce recruitment of tryptophan aspartate-containing coat protein (TACO), which associates with the phagosomal membrane in a cholesterol-dependent way and prevents its fusion with lysosomes (Gatfield and Pieters 2000a). A report by Wang and coworkers demonstrate that depletion of lipid rafts by methyl-β-cyclodextrin resulted in reduced survival and virulence of Porphyromonas gingivalis, a periodontal pathogen in the host. Interestingly, lipid rafts disruption was shown to promote the colocalization of internalized P. gingivalis with lysosome (Wang and Hajishengallis 2008). While Chlamydia trachomatis, a common sexually transmitted pathogen also use lipid rafts to disguise the phagosome as a vesicles which are derived from Golgi apparatus. These phagosomes are transported to Golgi apparatus to capture sphingolipid-rich vesicles to disguise host defense system (Norkin et al. 2001). These studies clearly indicate an important role of lipid rafts in pathogen survival beyond entry. Therefore, it is important to understand the mechanism(s) of invasion used by pathogens in order to design the successful strategy for regulating disease pathogenesis.
Mechanisms of bacterial entry in cells
Caveolae, Clathrin-coated pits and phagocytosis all provide entry pathways to toxins, and fall within the size range of 60-100 nm, respectively (Conner and Schmid 2003; Rollason et al. 2007). Of these, clathrin-coated pits are considered raft-free, while the caveolae pathway has been considered to be raft-mediated. Interestingly, Shiga toxin and the protective antibody against anthrax toxin both associate with rafts on the cell surface and are subsequently internalized via clathrin-coated pits (Kurzchalia 2003; Sandvig and van Deurs 2002a, b). Association of caveolae with E. coli-containing phagosomes has been reported during its cellular entry (Jermy 2010; Zaas et al. 2005), which reflects that caveolae may provide signaling platforms required for bacterial engulfment.
Signaling pathways regulated during bacterial infections
The entry of bacteria through lipid rafts leads to changes in signaling pathways including autophagy, alterations in inflammatory cytokine production, and induction of apoptosis (Fig. 1). Some bacteria like Salmonella typhimurium and S. flexneri, modulate host signaling by secreting bacterial invasion effector proteins which associate with membrane rafts (Manes et al. 2003b; Lafont et al. 2002; Lacalle et al. 2002). Further, while cellular entry of certain toxins and bacteria have been found to occur through raft-containing pathways, the step(s) required may differ depending on the pathogens.

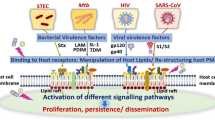
Role of caveolae and lipid rafts in bacterial entry. Bacteria usually enters the cell through endocytic pathway and survives by avoiding fusion with lysosome. The entry is mediated through caveolae or lipid rafts and can be targeted to a number of different intracellular compartments. Bacteria that enter in a lipid raft-dependent manner trigger signaling pathways including induction of apoptosis and cytokine production
Besides utilizing rafts to generate phagosomes as a survival mechanism within non-professional and professional phagocytic cells (Gatfield and Pieters 2000b; Kim et al. 2002; Ferrari et al. 1999; Watarai et al. 2001), bacteria like L. monocytogenes, an intracellular bacterium, have been shown to augment raft-mediated signaling thereby leading to increased cytokine production at the site of infection (Seveau et al. 2004; Coconnier et al. 2000; Parihar et al. 2013; Gekara et al. 2010; Barbuddhe and Chakraborty 2008; Kayal and Charbit 2006). Moreover, recent studies have shown impaired immune responses and reduced survival of caveolin-1-deficient mice in response to pulmonary challenge with Klebsiella pneumoniae and Pseudomonas aeruginosa (Guo et al. 2012; Gadjeva et al. 2010). Further, it was observed that knocking down caveolin-1 resulted in reduced bacterial clearance and an increase in production of pro-inflammatory cytokines in Klebsiella-challenged lung epithelial MLE-12 cells (Guo et al. 2012). Lipid rafts have thus been proposed to modulate bacterial internalization, which may impact host repair mechanisms, a critical phenomenon for host defense against the possibility of DNA damage caused by K. pneumoniae infection (Huang et al. 2013). It was reported that lungs from caveolin-1 deficient mice showed suppressed NF-кB activation in response to intraperitoneal injection of lipopolysaccharide, an effect believed to be mediated via endothelial NO synthase (eNOS) (Garrean et al. 2006) (Fig. 2). P. aeruginosa, on the contrary, induces the formation of ceramide-rich rafts, which not only promote host-pathogen interactions but also induces apoptosis of infected cells (Grassme et al. 2003a). Together, these studies suggest that lipid rafts play critical roles not only in bacterial entry, but also in influencing the host inflammatory responses critical for promoting or clearing infection.

Bacterial entry and interaction with receptor molecules. a Entry of bacteria into a host cell involves specific interactions of bacterial PAMPs (e.g., LPS and lipopeptides) with receptor molecules on cell surface, leading to conformational changes in rafts. b Upon stimulation by bacterial lipopeptides, TLR2 forms heterodimers with TLR6 in lipid raft domains, while CD14 and CD36 (cluster of differentiation) act as adaptor molecules for ligand transfer onto TLR2/6 heterodimers. Following ligand stimulation TIRAP binds to TLR2 through TIR domains and recruits adaptor protein MyD88, which then activates of TNF receptor associated factor 6 (TRAF6). This pathway induces the activation of NF-кB pathway leading to production and secretion of pro-inflammatory cytokines
Bacterial membrane microdomains
In addition to the role of host cell membrane domains during infections, bacterial proteins appear to recognize specific sites on their own membranes as well. Recently Lopez and Kolter described the existence of functional microdomains in bacterial membranes that harbor homologs of Flotillin-1 (eukaryotic lipid raft protein) and are rich in polyisoprenoids and other signaling and transport proteins (Lopez 2015; Lopez and Kolter 2010). Bacterial membrane proteins contained in lipid rafts function similarly to those found in eukaryotes and are involved in cell signaling pathways (Barak and Muchova 2013). Furthermore, as discussed by Barak and Muchova, differential aspects of a bacterial membrane’s physical characteristics, specifically, a negative curvature (concave) or a positive curvature (convex), are recognized by bacterial proteins. Specifically, membranes with a convex shape are linked with asymmetric cell division. Yet another physical characteristic that is important for the organization/localization of proteins in bacterial membranes is the transmembrane electric potential (Barak and Muchova 2013). As utilized for study of eukaryotic lipid rafts, the procedure used to analyze bacterial rafts is based on their ability to resist disaggregation by non-ionic detergents followed by separation by sucrose gradient. Studies conducted by Lopez and coworkers demonstrate that the detergent-resistant membranes (DRM) of S. aureus involves the proteins required for biofilm formation, virulence, and signaling (Lopez and Kolter 2010). Moreover, all of these processes can be inhibited by blocking the formation of lipid rafts with the use of small molecules (Bramkamp and Lopez 2015). Zaragozic acid (ZA), a known inhibitor of squalene synthase, which is involved in the formation of polyisoprenoids , inhibits the formation of lipid rafts resulting in impaired biofilm formation. In a recent study, Somani and coworkers demonstrated constitutive expression of FlotP, a Flotillin-1 homolog in Bacillus anthracis. The importance of rafts in the growth, membrane fluidity, and virulence of B. anthracis was demonstrated by studies conducted using ZA (Somani et al. 2016). Evidence of detergent soluble and detergent-resistant membrane fractions in Bacillus halodurans and Bacillus subtilis and the presence of flotillin-1 like protein (FloT) in the cytoplasmic membrane of the latter was also provided in recent studies (Donovan and Bramkamp 2009; Zhang et al. 2005). The membranes microdomains of bacterium Borrelia burgdorferi, which causes lime disease, have also been shown to possess properties similar to eukaryotic lipid rafts. Likewise, B. burgdorferi domains favor selective incorporation of molecules which are membrane anchored via saturated acyl chains but not those having unsaturated acyl chains (LaRocca et al. 2013). The other interesting observation reported in this study suggests that any changes in B. burgdorferi membrane morphology and integrity may affect the physical connection between flagella and the outer membranes. This may result in loss of flat wave morphology, which changes the membrane permeability leading to loss of flagellar subunits followed by death of spirochetes due to increased sensitivity to secondary osmotic lysis. These events confirm the functional importance of membrane microdomains in B. burgdorferi (LaRocca et al. 2013).
Bacterial membrane microdomains were discovered relatively recently, and therefore, increased understanding is still needed. Future studies need to focus on determining the structural components of bacterial rafts and their biological role in different bacterial species. Further, since the perturbation of bacterial microdomains can affect several physiological processes associated with pathogenesis, this field of study also may open new avenues for the control of bacterial infections.
Lipid rafts and viral infections
Viruses are classified into four basic types based on their outer structure. These include the following: enveloped DNA viruses, enveloped RNA viruses, non-enveloped DNA viruses, and non-enveloped RNA viruses. In addition, the general virus cycle of infection is typically divided into four stages: entry, translation, replication, and assembly/release (Fig. 3). The early stage of viral entry into the host cell involves the binding of the virus to one or more cell-surface receptors followed by entry into the cell (Ono and Freed 2005). Study of the localization of viral structural proteins and the effects of use of cholesterol/raft disrupting agents on the replication of several viruses have demonstrated the role of membrane microdomains in viral cycle.

Entry of virus through lipid rafts. Enveloped viruses enter the cells through direct fusion between the viral membrane and cell-surface receptor or through endocytic-mediated pathway, whereas in non-enveloped viruses are translocated directly into cytoplasm by destruction of the viral capsid. Transcription and replication process of viral genome in non-enveloped viruses takes place in the nucleus but for enveloped viruses it takes place both in nucleus and cytoplasm. After assembly the viral proteins, viral glycoprotein’s and infectious particles of viruses are released from cells via budding process at intracellular membranes (e.g., the ER) and at the plasma membrane which disrupts the humoral and cellular responses of the cell
Viruses generally enter cells through one of two basic mechanisms: (1) enveloped RNA viruses from the Rhabdoviridae, Flaviviridae, Orthomyxoviridae, and Togaviridae families and enveloped DNA viruses from the Papovaviridae and Adenoviridae families enter through endocytic vesicles and (2) through translocation or penetration of viral particles directly into the cytoplasm, by fusion of the viral envelope at the plasma membrane, or through destruction of the viral capsid as in the case of viruses from the Paramyxoviridae family (Takahashi and Suzuki 2011a). In contrast, viruses from the Herpesviridae family utilize both pathways (Takahashi and Suzuki 2011a). Moreover, most viral genomes of enveloped RNA viruses replicate and are transcribed in the cytosol, while those of enveloped DNA viruses replicate in the nucleus. Progeny viruses subsequently bud and are released from the cell membrane.
Involvement of surface receptors in viral entry
There is plethora of information regarding the involvement of cell-surface receptors localized within lipid rafts in facilitating viral entry. In brief, facilitation of HIV-1 entry into the cell by binding to the CD4 receptor causes conformational changes in the surface glycoprotein gp120 resulting in its interaction with CXCR4 or CCR5. These events lead to conformational changes in the transmembrane glycoprotein gp41 leading to unmasking of its fusogenic domain (Fantini et al. 1997; Hammache et al. 1998; Hug et al. 2000). While glycosyl phosphatidylinositol (GPI) anchored folate receptor-α (FRα) localized in membrane rafts is required for the entry of ebola and marburg viruses to enter host cells. The EBV human herpes virus infects B lymphocytes via the human complement receptor type 2 (CR2 aka CD21), which exists as a complex with CD19 and palmitoylated CD81 in lipid rafts. This CD19/21/81 complex functions to prolong the stability of, and signaling via, the B cell antigen receptor (Fingeroth et al. 1984; Cherukuri et al. 2004; Birkenbach et al. 1992). While discussion about all the related studies is beyond the scope of this article, most of the work in this area is referenced in Table 3.
Mechanisms of viral entry in cells
Following the attachment of non-enveloped viruses to cell-surface receptors, the bound virions can be internalized through a process mediated by rafts involving membrane invaginations and intracytoplasmic vesiculation (Ono and Freed 2005; Takahashi and Suzuki 2011a; Suzuki and Suzuki 2006). This type of viral entry, also termed atypical endocytosis, has been observed for the SV40 (Papovaviridae family) virus. Entry of SV40 into target cells is mediated through an association of major histocompatibility complex-1 (MHC-1) with caveolae (Stang et al. 1997; Anderson et al. 1998a; Norkin et al. 2002a; Pelkmans et al. 2001) or the GM1 ganglioside receptor in lipid rafts (Tsai et al. 2003a; Pelkmans 2005). Virus-incorporated caveolae then undergo budding, and the caveolae carries the virus to the endoplasmic reticulum (ER) along cellular microtubules.
Non-enveloped DNA viruses also enter into cells through typical endocytosis utilizing clathrin-dependent or clathrin-independent and caveolae/raft-dependent pathways followed by release of their viral DNA genomes into the cytoplasm by viral capsid destruction (Ono 2010). Reports suggest that Coxsackievirus, which causes aseptic meningitis with symptoms resembling a respiratory disease in addition to flaccid paralysis and chronic myocarditis, utilizes the cellular coxsackievirus and adenovirus receptor for its entry via membrane rafts (Triantafilou and Triantafilou 2003; 2004). In addition, several enteroviruses, which cause nerve paralysis, cerebral meningitis, anathema and respiratory symptoms, also utilize caveolae-dependent or raft-dependent endocytosis mechanisms for cellular entry (Stuart et al. 2002a; Marjomaki et al. 2002; Upla et al. 2004; Pietiainen et al. 2005). In contrast, entry of rhinovirus is mediated by ceramide-rich rafts (Grassme et al. 2005).
Enveloped viruses also use rafts during the internalization and fusion process. The entry of enveloped viruses involves virus attachment followed by close apposition of the virus and plasma membranes. The two membranes then fuse to deliver the virus’ genomic RNA into the host cells, which requires conversion of the virus-encoded envelope glycoprotein (Env) from its native state to its fusion-activated form (Manes et al. 2003a; Fantini et al. 2002; Chazal and Gerlier 2003; Brown et al. 2002a)
Virus assembly
The later stages of the virus infection cycle involve the assembly of viral components into virions and maturation into infectious particles. Viruses use lipid rafts as a platform because their structure offers an efficient system for concentrating all the viral proteins and glycoproteins required for the assembly of new virions (Ono and Freed 2005; Suzuki and Suzuki 2006; Das et al. 2010a). There is abundant evidence to support the idea that membrane lipids are not randomly incorporated into the viral envelope. Further, it has also been suggested that viral glycoproteins determine the site of virus assembly and budding (Vieira et al. 2010; Garoff and Simons 1974; Garoff et al. 1974; Allison et al. 1995; Vennema et al. 1996; Bruss 2004). Moreover, in polarized epithelial cells, the viral glycoproteins contain sorting signals or motifs and are directed to the specific site where assembly and budding will occur (Vieira et al. 2010; Nayak et al. 2004). Finally, the lipid composition of the influenza virus family is affected by the affinity of the haemagglutinin and neuraminidase glycoproteins for these lipids, while some reports also suggest that the influenza virus buds from raft domains (Vieira et al. 2010; Chazal and Gerlier 2003; Nayak et al. 2004). HIV-1 is also enclosed in a lipid envelope enriched in cholesterol and sphingolipids, suggesting specific membrane localization is required for assembly (Vieira et al. 2010; Aloia et al. 1993; Campbell et al. 2001; Raulin 2002). With similar methods of assembly and budding within membrane rafts, many other viruses, including influenza virus, measles virus, Ebola virus, and possibly Sendai virus, also use lipid rafts as assembly platforms (Vieira et al. 2010; Luo et al. 2008). In this regard, it was also suggested that the RSV (respiratory syncytial virus) assembles within lipid rafts where viral proteins colocalize with caveolin-1 (Vieira et al. 2010; Brown et al. 2002a, b). Taken together, these structures then have the ability to regulate budding; however, the mechanism by which the lipid raft can favor the budding and/or fission process has yet to be explored in detail (Vieira et al. 2010; Nayak et al. 2004; Luo et al. 2008).
For enveloped viruses, the infectious particles are released from the cell via the budding process. Furthermore, the budding of new virions from the raft allows the exclusion or inclusion of specific host cell membrane proteins in the virus particle, which can disrupt cellular and/or humoral immune responses to the virus (Suzuki and Suzuki 2006; Das et al. 2010a). Non-enveloped viruses are released from the infected cell by disrupting the plasma membrane, whereas enveloped viruses contain a host cell-derived lipid bilayer, which is acquired during budding. Only cytolytic viruses lead to disruption of cell integrity, while other viruses are released in a non-destructive manner. In this context, studies demonstrated that ebola and marburg viruses use viral glycoprotein-bearing rafts for budding from host cells. Specifically, VP40 (ebola virus matrix protein) oligomerizes with rafts and plays an important role in assembly and budding(Bavari et al. 2002a), while glycoprotein complexes composed of gp120 and gp41 along with membrane rafts are used for HIV-1 assembly (Bhattacharya et al. 2004; Ono et al. 2005; Ono et al. 2007; Leung et al. 2008a). Koshizuka’s group demonstrated the association of HSV-1 UL11 (protein involved in virion maturation) with lipid rafts (Koshizuka et al. 2007). Among non-enveloped viruses, the association of viral protein VP4 and NSP4-purified virions with lipid rafts have been studied for rotavirus (Sapin et al. 2002; Cuadras and Greenberg 2003) while an outer capsid protein important for virus assembly, VP5, was observed to co-purify with lipid raft entities during bluetongue virus infection. The association of VP5 with rafts was shown to depend on membrane docking domain similar to what occurs with the soluble N-ethylmaleimide-sensitive fusion attachment protein receptor (SNARE) family (Bhattacharya and Roy 2008a). Although rafts are involved in virus assembly, we have to keep in mind that only a fraction of viral proteins are found associated with rafts. However, this could be due to the poor biochemical characterization of raft subsets or to the transient nature of the association (Vieira et al. 2010).
Signaling pathways regulated during viral infections
Since membrane rafts are involved in the entry, assembly, and budding processes of several viruses, it is not surprising that cellular signaling pathways are also affected during viral infections. Lee et al. reported the anti-apoptotic role of PI3K signaling in response to dengue and japanese encephalitis virus (JEV) infections in human lung carcinoma and murine neuroblastoma cell lines (Lee et al. 2005). However, using microglial cells, Chen and coworkers demonstrated the induction of the Src/Ras±/Raf/ERK/NF-кB pathways leading to induction of TNF-α and IL-1β in response to challenge with JEV (Chen et al. 2011). This is important because Src-mediated signaling induced by JEV is known to play a critical role in neuronal cell death. Earlier studies revealed the role of lipid rafts in the activation of Src, Ras, Raf, ERK, and NF-кB, which all contribute to JEV-induced TNF-α and IL-1β production (McCubrey et al. 2007; Wu et al. 2015). Furthermore, involvement of lipid rafts in Src-mediated signaling pathways upstream of the activation of Raf/ERK/NF-кB has been reported in response to several stimuli (Roux and Blenis 2004). These studies provide substantial evidence that lipid rafts play an important role in Src/Ras/Raf/ERK/ NF-кB signaling during JEV infection, thereby influencing host immune responses. In addition, Bentley and coworkers (Bentley et al. 2007a) also demonstrated that rhinovirus (RV) activates PI3K and Akt, which leads to IL-8 expression. Further, they observed that RV colocalizes with Src, PI3K, and Akt in membrane rafts. These and several related findings demonstrate that lipid raft-mediated signaling pathways are regulated during viral infections and play critical role in disease pathogenesis (Bentley et al. 2007a; Silva et al. 2011; Ebihara et al. 2008; Huang et al. 2009b; Fang et al. 2010) (Fig. 2).
Lipid rafts and fungal infections
Information regarding the role of lipid rafts in fungal infections is evolving; however, initial studies suggest that rafts facilitate fungal infections by regulating hyphal growth and biofilm formation, and by concentrating virulance factors (VFs) (Vieira et al. 2010; Farnoud et al. 2015).
Role of the host rafts during fungal infection and modulation of host signaling
The connection of fungal infection with lipid rafts has not been extensively explored; however, evidences for modulation in host cell signaling pathways by mycopathogens has been demonstrated. Paracoccidioidomycosis is one of common fungal disease with high prevalence in South and Central America and is caused by Paracoccidioides brasiliensis. Infection with P. brasiliensis promotes the aggregation of lipid rafts in epithelial cells, which then support fungal adhesion and activation of Src kinases (Fig. 4). Localization of Ganglioside (GM1) in the host membrane microdomains allows P. brasiliensis-epithelial cell contact and fugal adhesion, which was found to be inhibited by methyl-β-cyclodextrin (MβCD) (Maza et al. 2008). Moreover, invasion by Candida albicans or P. brasiliensis leads to the activation of host cell tyrosine kinases which can be regulated by tyrosine kinase inhibitors (Belanger et al. 2002; Monteiro da Silva et al. 2007). Further, hyphal growth and biofilm formation facilitate invasive growth of Cryptococcus neoformans into the central nervous system; for which the pathogen needs to travel across the blood-brain barrier composed of human brain microvascular endothelial cells (HBMECs). CD44 is the C. neoformans receptor believed to be anchored in lipid rafts on the surface of HBMECs during infection. It is known that in caveolin-1 knockdown HBMECs, GM1 and CD44 protein become randomly distributed and clustered into irregular-islands on the cell surface (Long et al. 2012). This suggests an important structural role of caveolin-1 in C. neoformans invasion. Activated PKCα, can directly interact with caveolin-1, on the plasma membrane, and is required for C. neoformans invasion into HBMEC (Jong et al. 2008; Oka et al. 1997). Finally, the microsporidiam parasite, Encephalitozoon cuniculi, which affects the nervous system, as well as the respiratory and digestive tracts, has been shown to localize to lipid rafts. While residing in a parasitophorous vacuole, this pathogen is surrounded by host cell lipids; which could be labeled with DilC16 (1,1′-dihexadecyl-3,3,3′,3′-tetramethylindocarbocyanine), a marker for lipid rafts, and DiO (3,3′-dilinoleyloxacarbocyanine), a marker for non-raft membrane domains. These results suggest that rafts contribute to the formation of parasitophorous vacuole membrane (Ronnebaumer et al. 2008) and also that mycopathogens may also infect cells in a raft-dependent manner (Vieira et al. 2010).

Fungal infection in association with lipid rafts and host cell signaling. Fungal infection promotes the aggregation of lipid rafts in epithelial cells. Localization of ganglioside (GM1) in the host membrane microdomains allows cell contact and fugal adhesion. Fungal adhesion leads to activation of PKCα and Src protein tyrosine kinases leading to downstream signaling events in the host cells. Lipid rafts facilitate fungal infections by regulating hyphal growth, biofilm formation, concentrating virulance factors, and drug resistance
Role of fungal rafts in virulence and drug resistance
The primary raft-forming lipids in C. albicans, a dimorphic opportunistic pathogen, include ergosterol and glycosphingolipids, while the transporter of N-acetylglucosamine protein (Ngt1), an efflux pump and drug transporter protein (CaCdr1p), GPI-anchored protein (Gas1p), and proton pump protein (Pma 1) are the major proteins associated with these rafts (Martin and Konopka 2004; Simonetti et al. 1974; Pasrija et al. 2008). C. albicans rafts are capable of forming biofilms creating a protected niche for microorganism against antibiotics and a source of persistent infection. The efficiency of biofilm formation can be significantly reduced after treatment with myriocin and aureobacidin suggesting an important role of sphingolipids (lipid rafts) in biofilms formation (Martin and Konopka 2004; Lattif et al. 2011). Furthermore, displacement of these proteins from membrane by disruption of ergosterol or the sphingolipid biosynthesis pathway negatively influences the drug resistance of fungi and suggests the importance of raft assembly for localization of these proteins (Wang et al. 2012). Similarly, when CaCdr1p is depleted from DRMs, Saccharomyces cerevisiae becomes prone to methotrexate and fluconazole (Pasrija et al. 2008). Furthermore, ergosterol depletion has also been shown to impair phagocytosis of Histoplama capsulatum, which may result from the release of proteins with anti-phagocytic function (Tagliari et al. 2012). Although, secretion of anti-phagocytic proteins has been demonstrated in case of C. neoformans, the direct link with ergosterol depletion and secretion of these proteins during H. capsulatum infection has not yet been established (Tagliari et al. 2012; Luberto et al. 2003). While studying chitosan-resistant and chitosan-sensitive filamentous fungi, Palma-Guerrero and coworkers demonstrated the importance of fungal rafts in drug resistance. They identified an Neisseria crassa desaturase mutant with chitosan resistance and high membrane rigidity. The polymeric compound chitosan exerts its fungicial effect by permeabilizing plasma membrane of filamentous fungi (Palma-Guerrero et al. 2010). Additionally, the role of detergent-resistant membranes in fungi for concentrating VFs-phospholipase B1 and Cu/Zn superoxide dismutase; and proton pump protein Pma1 was demonstrated by Siafakas et al. and Holyoak groups, respectively (Siafakas et al. 2006; Holyoak et al. 1996). Several reports have demonstrated the functional role of these VFs in evading hosts immune responses and Pma1 in the pathogenicity of C. neoformans (Farnoud et al. 2014).
Perspectives
There is abundance of literature defining the role of lipid rafts in several pathological conditions including Alzheimer’s disease, Prion disease (Campbell et al. 2001; Peters et al. 2003; Liao et al. 2001a; Nguyen and Taub 2002a; Liao et al. 2003), abnormalities associated with the cardiovascular system such as hypertension and cardiac hypertrophy (Fantini et al. 2002; Peters et al. 2003; Liao et al. 2001a; Golde and Eckman 2001) and during bacterial and viral infections. Therefore, understanding the regulation and formation/composition of rafts is imperative for the design of therapeutic strategies for prevention and cure of related pathologies, as well as to aid in patient management. For example, the use of statins in cancer chemotherapy has been suggested as these drugs have been shown to alter endothelial cell function and inflammatory responses (Mo and Elson 2004). However, the effect of statins is not directly mediated through cholesterol, but is rather through their effect on isoprenoids, which share initial biochemical synthetic steps with cholesterol and play important roles in anchoring several GTPases in the membrane (Vaughan 2003). Similar effects including the activation of GTPase were reported in cholesterol depletion and repletion models used to study the raft function in cell culture models (Pierini et al. 2003; Nagao et al. 2007). In this regard, a membrane-tethering inhibitor of β-secretase that targets endosomes was designed for treatment of Alzheimer’s disease (Rajendran et al. 2008). This same idea may be employed for development of small lipophilic inhibitors to target raft-anchored proteins that are activated in membrane compartments.
Earlier reports have also demonstrated that modulating the structure/composition of rafts can lead to inhibition of the infection cycle of HIV-1, which enters the host cell through rafts, and can regulate raft-mediated signaling pathways. SP01A, a widely used inhibitor of HIV-1 infection, affects cholesterol synthesis and regulates the cholesterol content of rafts thereby preventing HIV-1 fusion with CD4+ cells. Likewise, the use of anti-raft drugs shows clear promise as an alternative to antibiotics to help reduce the threat of drug-resistant bacteria. Further, targeting lipid raft formation in bacteria using small molecules has also been shown to reduce their virulence by regulating diverse processes including exoprotease production, biofilm formation, and attachment (Kong et al. 2006; Liu et al. 2008). However, careful assessment of the strategies utilized should be made while targeting cholesterol in the rodent models, as there exists quantitative and qualitative difference in its balance as compared to humans. Further, high concentrations of cholesterol at several anatomical sites would also impair the ability to uniformly target cholesterol for pathological conditions that effect various tissues/organs (Dietschy and Turley 2002).
Given that there is a unique assembly of proteins and lipids during the formation of signaling/raft domains in response to infection with various pathogens, alteration of the composition of lipid rafts through nutritional intervention may be other interesting approach to combat pathogens and modulate the inflammatory responses. This strategy has received wide acceptance in the recent years and growing evidence suggests that consumption of n-3 long-chain polyunsaturated fatty acids (LCPUFAs) can reduce the risk of cardiovascular disease and breast cancer. In this context, n-3 LCPUFA has been shown to alter the partitioning of proteins and composition of rafts including the reduction of cholesterol (Hashimoto et al. 1999) and caveolae (Ma et al. 2004), and has been shown to regulate Akt and NF-кB activation(Schley et al. 2007). In addition, n-3 LCPUFA induces p38MAPK and EGFR phosphorylation, which leads to increased apoptosis in tumor cells (Schley et al. 2007; Tikhomirov and Carpenter 2004). An extensive report was published by Turk and Chapkin (Turk and Chapkin 2013) describing the regulation of lipid raft size, composition, and associated signaling mechanisms by n-3 LCPUFA. These findings underscore the need to further assess the therapeutic potential of n-3 LCPUFA in disease states mediated by lipid raft-associated processes.
Conclusions
With the majority of the scientific community focusing on DNA, RNA, and proteins to understand the pathogenesis of disease, the role of lipids has remained relatively unexplored. However, recent advances in the field of membrane microdomains/lipid rafts research has opened new avenues to investigate alternative therapeutic intervention strategies for many diseases. Although there is abundant literature suggesting important role of lipid rafts in bacterial, viral, and fungal infections, several questions still remain to be answered in order to define and target raft-mediated responses. Thus, impairment of bacterial and viral entry and propagation in the host in response to deviations/alterations in functional membrane microdomains warrants further study in order to better assess their role in bacterial and viral infections. Further, the involvement of structural components in terms of regulating rafts size, the concentration of lipids in microdomains, the specificity of protein and lipid clustering, and rafts scaffolding during disease pathogenesis are areas requiring additional focus and potentially, even more sophisticated techniques. Addressing these particular aspects of functional membrane microdomains will lead to a better understanding of disease mechanisms and potentially more effective treatment strategies including cell-based or personalized medicine approaches. The former approach is based on the concept of delivering therapeutic agents/drugs to specific cell types using nano-particle based delivery of drugs to the plasma membrane of cells for raft-dependent intracellular trafficking (Partlow et al. 2008). With regard to personalized medicine approaches, lipidomics monitoring can provide a description of the concentration, structure, and function of complete sets of lipids as well as information regarding the interaction of lipids with proteins and other metabolites. This information can be utilized to design a personalized strategy to restore membrane balance using lipids and cofactors from nutritional sources (Escriba 2006). In addition, recent reports have suggested increased individual disease susceptibility to several diseases due to the limitations of several therapeutic proteins like growth factors and antibodies to reach the cytoplasm of diseased cells. These limitations can be overcome through covalent modification with glycolipid structures (GPI anchors) resulting in directed lipid raft-mediated signaling. Furthermore, research and clinical studies using anticancer drugs that target lipid messengers or their carriers are ongoing. These drugs are designed to disrupt crosstalk between cells within the tumor microenvironment and cancer cells (Muller 2011). Future studies aiming to reduce the generation and secretion of exosomes, which possess the bioactive lipids found in the extracellular space in tumor environment, have also been proposed by the experts in cancer research. These new approaches hold promise and the potential to offer novel therapeutic targets with translation potential for future intervention studies.
References
Akhtar J, Tiwari V, Oh M-J, Kovacs M, Jani A, Kovacs SK, Valyi-Nagy T, Shukla D. HVEM and nectin-1 are the major mediators of herpes simplex virus 1 (HSV-1) entry into human conjunctival epithelium. Invest Ophthalmol Vis Sci. 2008;49:4026–35.
Akula SM, Pramod NP, Wang F-Z, Chandran B. Integrin α3β1 (CD 49c/29) is a cellular receptor for Kaposi's sarcoma-associated herpesvirus (KSHV/HHV-8) entry into the target cells. Cell. 2002;108:407–19.
Alfsen A, Yu H, Magérus-Chatinet A, Schmitt A, Bomsel M. HIV-1-infected blood mononuclear cells form an integrin-and agrin-dependent viral synapse to induce efficient HIV-1 transcytosis across epithelial cell monolayer. Mol Biol Cell. 2005;16:4267–79.
Allison SL, Schalich J, Stiasny K, Mandl CW, Kunz C, Heinz FX. Oligomeric rearrangement of tick-borne encephalitis virus envelope proteins induced by an acidic pH. J Virol. 1995;69:695–700.
Aloia RC, Tian H, Jensen FC. Lipid composition and fluidity of the human immunodeficiency virus envelope and host cell plasma membranes. Proc Natl Acad Sci U S A. 1993;90:5181–5.
Alonso MA, Millan J. The role of lipid rafts in signalling and membrane trafficking in T lymphocytes. J Cell Sci. 2001;114:3957–65.
Altelaar AF, Klinkert I, Jalink K, de Lange RP, Adan RA, Heeren RM, Piersma SR. Gold-enhanced biomolecular surface imaging of cells and tissue by SIMS and MALDI mass spectrometry. Anal Chem. 2006;78:734–42.
Ammendolia MG, Bertuccini L, Minelli F, Meschini S, Baldassarri L. A Sphingomonas bacterium interacting with epithelial cells. Res Microbiol. 2004;155:636–46.
Anderson RG. The caveolae membrane system. Annu Rev Biochem. 1998;67:199–225.
Anderson HA, Chen Y, Norkin LC. MHC class I molecules are enriched in caveolae but do not enter with simian virus 40. J Gen Virol. 1998a;79(Pt 6):1469–77.
Anderson HA, Chen Y, Norkin LC. MHC class I molecules are enriched in caveolae but do not enter with simian virus 40. J Gen Virol. 1998b;79:1469–77.
Androutsopoulos G, Thanatsis N, Michail G, Adonakis G, Decavalas G. Initial steps and mechanisms of HPV infection. HJOG. 2015;14.
Anes E, Kuhnel MP, Bos E, Moniz-Pereira J, Habermann A, Griffiths G. Selected lipids activate phagosome actin assembly and maturation resulting in killing of pathogenic mycobacteria. Nat Cell Biol. 2003;5:793–802.
Arana L, Gangoiti P, Ouro A, Trueba M, Gomez-Munoz A. Ceramide and ceramide 1-phosphate in health and disease. Lipids Health Dis. 2010;9:15.
Arruda E, Crump C, Hayden F. In vitro selection of human rhinovirus relatively resistant to soluble intercellular adhesion molecule-1. Antimicrob Agents Chemother. 1994;38:66–70.
Bacia K, Scherfeld D, Kahya N, Schwille P. Fluorescence correlation spectroscopy relates rafts in model and native membranes. Biophys J. 2004;87:1034–43.
Barak I, Muchova K. The role of lipid domains in bacterial cell processes. Int J Mol Sci. 2013;14:4050–65.
Barbuddhe S, Chakraborty T. Biotechnological applications of Listeria's sophisticated infection strategies. Microb Biotechnol. 2008;1:361–72.
Bauer M, Pelkmans L. A new paradigm for membrane-organizing and -shaping scaffolds. FEBS Lett. 2006;580:5559–64.
Bavari S, Bosio CM, Wiegand E, Ruthel G, Will AB, Geisbert TW, Hevey M, Schmaljohn C, Schmaljohn A, Aman MJ. Lipid raft microdomains: a gateway for compartmentalized trafficking of Ebola and Marburg viruses. J Exp Med. 2002a;195:593–602.
Bavari S, Bosio CM, Wiegand E, Ruthel G, Will AB, Geisbert TW, Hevey M, Schmaljohn C, Schmaljohn A, Aman MJ. Lipid raft microdomains a gateway for compartmentalized trafficking of Ebola and Marburg viruses. J Exp Med. 2002b;195:593–602.
Belanger PH, Johnston DA, Fratti RA, Zhang M, Filler SG. Endocytosis of Candida albicans by vascular endothelial cells is associated with tyrosine phosphorylation of specific host cell proteins. Cell Microbiol. 2002;4:805–12.
Bentley JK, Newcomb DC, Goldsmith AM, Jia Y, Sajjan US, Hershenson MB. Rhinovirus activates interleukin-8 expression via a Src/p110beta phosphatidylinositol 3-kinase/Akt pathway in human airway epithelial cells. J Virol. 2007a;81:1186–94.
Bentley JK, Newcomb DC, Goldsmith AM, Jia Y, Sajjan US, Hershenson MB. Rhinovirus activates interleukin-8 expression via a Src/p110β phosphatidylinositol 3-kinase/Akt pathway in human airway epithelial cells. J Virol. 2007b;81:1186–94.
Betz RC, Schoser BG, Kasper D, Ricker K, Ramirez A, Stein V, Torbergsen T, Lee YA, Nothen MM, Wienker TF, Malin JP, Propping P, Reis A, Mortier W, Jentsch TJ, Vorgerd M, Kubisch C. Mutations in CAV3 cause mechanical hyperirritability of skeletal muscle in rippling muscle disease. Nat Genet. 2001;28:218–9.
Bhattacharya B, Roy P. Bluetongue virus outer capsid protein VP5 interacts with membrane lipid rafts via a SNARE domain. J Virol. 2008a;82:10600–12.
Bhattacharya B, Roy P. Bluetongue virus outer capsid protein VP5 interacts with membrane lipid rafts via a SNARE domain. J Virol. 2008b;82:10600–12.
Bhattacharya J, Peters PJ, Clapham PR. Human immunodeficiency virus type 1 envelope glycoproteins that lack cytoplasmic domain cysteines: impact on association with membrane lipid rafts and incorporation onto budding virus particles. J Virol. 2004;78:5500–6.
Birkenbach M, Tong X, Bradbury LE, Tedder TF, Kieff E. Characterization of an Epstein-Barr virus receptor on human epithelial cells. J Exp Med. 1992;176:1405–14.
Blixenkrone-Møller M, Bernard A, Bencsik A, Sixt N, Diamond LE, Logan JS, Wild TF. Role of CD46 in measles virus infection in CD46 transgenic mice. Virology. 1998;249:238–48.
Bollinger CR, Teichgraber V, Gulbins E. Ceramide-enriched membrane domains. Biochim Biophys Acta. 2005;1746:284–94.
Bramkamp M, Lopez D. Exploring the existence of lipid rafts in bacteria. Microbiol Mol Biol Rev. 2015;79:81–100.
Brown DA. Lipid rafts, detergent-resistant membranes, and raft targeting signals. Physiology (Bethesda). 2006;21:430–9.
Brown G, Aitken J, Rixon HW, Sugrue RJ. Caveolin-1 is incorporated into mature respiratory syncytial virus particles during virus assembly on the surface of virus-infected cells. J Gen Virol. 2002b;83:611–21.
Brown G, Rixon HW, Sugrue RJ. Respiratory syncytial virus assembly occurs in GM1-rich regions of the host-cell membrane and alters the cellular distribution of tyrosine phosphorylated caveolin-1. J Gen Virol. 2002a;83:1841–50.
Brown G, Rixon HWM, Sugrue RJ. Respiratory syncytial virus assembly occurs in GM1-rich regions of the host-cell membrane and alters the cellular distribution of tyrosine phosphorylated caveolin-1. J Gen Virol. 2002c;83:1841–50.
Bruss V. Envelopment of the hepatitis B virus nucleocapsid. Virus Res. 2004;106:199–209.
Caliceti C, Zambonin L, Prata C, Vieceli Dalla Sega F, Hakim G, Hrelia S, Fiorentini D. Effect of plasma membrane cholesterol depletion on glucose transport regulation in leukemia cells. PLoS One. 2012;7:e41246.
Campbell SM, Crowe SM, Mak J. Lipid rafts and HIV-1: from viral entry to assembly of progeny virions. J Clin Virol. 2001;22:217–27.
Castro BM, Silva LC, Fedorov A, de Almeida RF, Prieto M. Cholesterol-rich fluid membranes solubilize ceramide domains: implications for the structure and dynamics of mammalian intracellular and plasma membranes. J Biol Chem. 2009;284:22978–87.
Chan SY, Empig CJ, Welte FJ, Speck RF, Schmaljohn A, Kreisberg JF, Goldsmith MA. Folate receptor-α is a cofactor for cellular entry by Marburg and Ebola viruses. Cell. 2001;106:117–26.
Chazal N, Gerlier D. Virus entry, assembly, budding, and membrane rafts. Microbiol Mol Biol Rev. 2003;67:226–37. table of contents
Chen CJ, Ou YC, Chang CY, Pan HC, Lin SY, Liao SL, Raung SL, Chen SY, Chang CJ. Src signaling involvement in Japanese encephalitis virus-induced cytokine production in microglia. Neurochem Int. 2011;58:924–33.
Cheong KH, Zacchetti D, Schneeberger EE, Simons K. VIP17/MAL, a lipid raft-associated protein, is involved in apical transport in MDCK cells. Proc Natl Acad Sci. 1999;96:6241–8.
Cherukuri A, Carter RH, Brooks S, Bornmann W, Finn R, Dowd CS, Pierce SK. B cell signaling is regulated by induced palmitoylation of CD81. J Biol Chem. 2004;279:31973–82.
Cho N-H, Kingston D, Chang H, Kwon E-K, Kim J-M, Lee JH, Chu H, Choi M-S, Kim I-S, Jung JU. Association of herpesvirus saimiri tip with lipid raft is essential for downregulation of T-cell receptor and CD4 coreceptor. J Virol. 2006;80:108–18.
Chung C-S, Huang C-Y, Chang W. Vaccinia virus penetration requires cholesterol and results in specific viral envelope proteins associated with lipid rafts. J Virol. 2005;79:1623–34.
Coconnier MH, Lorrot M, Barbat A, Laboisse C, Servin AL. Listeriolysin O-induced stimulation of mucin exocytosis in polarized intestinal mucin-secreting cells: evidence for toxin recognition of membrane-associated lipids and subsequent toxin internalization through caveolae. Cell Microbiol. 2000;2:487–504.
Cohen AW, Park DS, Woodman SE, Williams TM, Chandra M, Shirani J, Pereira de Souza A, Kitsis RN, Russell RG, Weiss LM, Tang B, Jelicks LA, Factor SM, Shtutin V, Tanowitz HB, Lisanti MP. Caveolin-1 null mice develop cardiac hypertrophy with hyperactivation of p42/44 MAP kinase in cardiac fibroblasts. Am J Physiol Cell Physiol. 2003;284:C457–74.
Conner SD, Schmid SL. Regulated portals of entry into the cell. Nature. 2003;422:37–44.
Cuadras MA, Greenberg HB. Rotavirus infectious particles use lipid rafts during replication for transport to the cell surface in vitro and in vivo. Virology. 2003;313:308–21.
Damm E-M, Pelkmans L, Kartenbeck J, Mezzacasa A, Kurzchalia T, Helenius A. Clathrin-and caveolin-1-independent endocytosis entry of simian virus 40 into cells devoid of caveolae. J Cell Biol. 2005;168:477–88.
Das S, Chakraborty S, Basu A. Critical role of lipid rafts in virus entry and activation of phosphoinositide 3' kinase/Akt signaling during early stages of Japanese encephalitis virus infection in neural stem/progenitor cells. J Neurochem. 2010a;115:537–49.
Das S, Chakraborty S, Basu A. Critical role of lipid rafts in virus entry and activation of phosphoinositide 3′ kinase/Akt signaling during early stages of Japanese encephalitis virus infection in neural stem/progenitor cells. J Neurochem. 2010b;115:537–49.
Deng D, Jiang N, Hao SJ, Sun H, Zhang GJ. Loss of membrane cholesterol influences lysosomal permeability to potassium ions and protons. Biochim Biophys Acta. 2009;1788:470–6.
Devaux PF, Herrmann A, Ohlwein N, Kozlov MM. How lipid flippases can modulate membrane structure. Biochim Biophys Acta. 2008;1778:1591–600.
Dhiman N, Jacobson RM, Poland GA. Measles virus receptors: SLAM and CD46. Rev Med Virol. 2004;14:217–29.
Dietschy JM, Turley SD. Control of cholesterol turnover in the mouse. J Biol Chem. 2002;277:3801–4.
DiNitto JP, Cronin TC, Lambright DG. Membrane recognition and targeting by lipid-binding domains. Sci STKE. 2003;2003:re16.
Dodes Traian MM, Gonzalez Flecha FL, Levi V. Imaging lipid lateral organization in membranes with C-laurdan in a confocal microscope. J Lipid Res. 2012;53:609–16.
Donovan C, Bramkamp M. Characterization and subcellular localization of a bacterial flotillin homologue. Microbiology. 2009;155:1786–99.
Doughty DM, Dieterle M, Sessions AL, Fischer WW, Newman DK. Probing the subcellular localization of hopanoid lipids in bacteria using NanoSIMS. PLoS One. 2014;9:e84455.
Drab M, Verkade P, Elger M, Kasper M, Lohn M, Lauterbach B, Menne J, Lindschau C, Mende F, Luft FC, Schedl A, Haller H, Kurzchalia TV. Loss of caveolae, vascular dysfunction, and pulmonary defects in caveolin-1 gene-disrupted mice. Science. 2001;293:2449–52.
Dragic T, Litwin V, Allaway GP, Martin SR, Huang Y, Nagashima KA, Cayanan C, Maddon PJ, Koup RA, Moore JP. HIV-1 entry into CD4+ cells is mediated by the chemokine receptor CC-CKR-5. 1996.
Ebihara T, Shingai M, Matsumoto M, Wakita T, Seya T. Hepatitis C virus-infected hepatocytes extrinsically modulate dendritic cell maturation to activate T cells and natural killer cells. Hepatology. 2008;48:48–58.
Eggeling C, Willig KI, Barrantes FJ. STED microscopy of living cells--new frontiers in membrane and neurobiology. J Neurochem. 2013;126:203–12.
Engelman DM. Membranes are more mosaic than fluid. Nature. 2005;438:578–80.
Escriba PV. Membrane-lipid therapy: a new approach in molecular medicine. Trends Mol Med. 2006;12:34–43.
Esen M, Schreiner B, Jendrossek V, Lang F, Fassbender K, Grassme H, Gulbins E. Mechanisms of Staphylococcus aureus induced apoptosis of human endothelial cells. Apoptosis. 2001;6:431–9.
Fang YT, Lin CF, Liao PC, Kuo YM, Wang S, Yeh TM, Shieh CC, Su IJ, Lei HY, Lin YS. Annexin A2 on lung epithelial cell surface is recognized by severe acute respiratory syndrome-associated coronavirus spike domain 2 antibodies. Mol Immunol. 2010;47:1000–9.
Fantini J, Garmy N, Mahfoud R, Yahi N. Lipid rafts: structure, function and role in HIV, Alzheimer's and prion diseases. Expert Rev Mol Med. 2002;4:1–22.
Fantini J, Hammache D, Delezay O, Yahi N, Andre-Barres C, Rico-Lattes I, Lattes A. Synthetic soluble analogs of galactosylceramide (GalCer) bind to the V3 domain of HIV-1 gp120 and inhibit HIV-1-induced fusion and entry. J Biol Chem. 1997;272:7245–52.
Farnoud AM, Mor V, Singh A, Del Poeta M. Inositol phosphosphingolipid phospholipase C1 regulates plasma membrane ATPase (Pma1) stability in Cryptococcus neoformans. FEBS Lett. 2014;588:3932–8.
Farnoud AM, Toledo AM, Konopka JB, Del Poeta M, London E. Raft-like membrane domains in pathogenic microorganisms. Curr Top Membr. 2015;75:233–68.
Ferrari G, Langen H, Naito M, Pieters J. A coat protein on phagosomes involved in the intracellular survival of mycobacteria. Cell. 1999;97:435–47.
Fingeroth JD, Weis JJ, Tedder TF, Strominger JL, Biro PA, Fearon DT. Epstein-Barr virus receptor of human B lymphocytes is the C3d receptor CR2. Proc Natl Acad Sci U S A. 1984;81:4510–4.
Gadjeva M, Paradis-Bleau C, Priebe GP, Fichorova R, Pier GB. Caveolin-1 modifies the immunity to Pseudomonas aeruginosa. J Immunol. 2010;184:296–302.
Gambin Y, Ariotti N, McMahon KA, Bastiani M, Sierecki E, Kovtun O, Polinkovsky ME, Magenau A, Jung W, Okano S, Zhou Y, Leneva N, Mureev S, Johnston W, Gaus K, Hancock JF, Collins BM, Alexandrov K, Parton RG. Single-molecule analysis reveals self assembly and nanoscale segregation of two distinct cavin subcomplexes on caveolae. elife. 2014;3:e01434.
Garcia-Marcos M, Pochet S, Tandel S, Fontanils U, Astigarraga E, Fernandez-Gonzalez JA, Kumps A, Marino A, Dehaye JP. Characterization and comparison of raft-like membranes isolated by two different methods from rat submandibular gland cells. Biochim Biophys Acta. 2006;1758:796–806.
Garoff H, Simons K. Location of the spike glycoproteins in the Semliki Forest virus membrane. Proc Natl Acad Sci U S A. 1974;71:3988–92.
Garoff H, Simons K, Renkonen O. Isolation and characterization of the membrane proteins of Semliki Forest virus. Virology. 1974;61:493–504.
Garrean S, Gao XP, Brovkovych V, Shimizu J, Zhao YY, Vogel SM, Malik AB. Caveolin-1 regulates NF-kappaB activation and lung inflammatory response to sepsis induced by lipopolysaccharide. J Immunol. 2006;177:4853–60.
Gatfield J, Pieters J. Essential role for cholesterol in entry of mycobacteria into macrophages. Science. 2000a;288:1647–51.
Gatfield J, Pieters J. Essential role for cholesterol in entry of mycobacteria into macrophages. Science. 2000b;288:1647–50.
Gault CR, Obeid LM, Hannun YA. An overview of sphingolipid metabolism: from synthesis to breakdown. Adv Exp Med Biol. 2010;688:1–23.
Gekara NO, Zietara N, Geffers R, Weiss S. Listeria monocytogenes induces T cell receptor unresponsiveness through pore-forming toxin listeriolysin O. J Infect Dis. 2010;202:1698–707.
Giri B, Dixit VD, Ghosh MC, Collins GD, Khan IU, Madara K, Weeraratna AT, Taub DD. CXCL12-induced partitioning of flotillin-1 with lipid rafts plays a role in CXCR4 function. Eur J Immunol. 2007;37:2104–16.
Goggel R, Winoto-Morbach S, Vielhaber G, Imai Y, Lindner K, Brade L, Brade H, Ehlers S, Slutsky AS, Schutze S, Gulbins E, Uhlig S. PAF-mediated pulmonary edema: a new role for acid sphingomyelinase and ceramide. Nat Med. 2004;10:155–60.
Golde TE, Eckman CB. Cholesterol modulation as an emerging strategy for the treatment of Alzheimer's disease. Drug Discov Today. 2001;6:1049–55.
Grassme H, Jendrossek V, Riehle A, von Kurthy G, Berger J, Schwarz H, Weller M, Kolesnick R, Gulbins E. Host defense against Pseudomonas aeruginosa requires ceramide-rich membrane rafts. Nat Med. 2003a;9:322–30.
Grassme H, Jendrossek V, Riehle A, Von Kürthy G, Berger J, Schwarz H, Weller M, Kolesnick R, Gulbins E. Host defense against Pseudomonas aeruginosa requires ceramide-rich membrane rafts. Nat Med. 2003b;9:322–30.
Grassmé H, Gulbins E, Brenner B, Ferlinz K, Sandhoff K, Harzer K, Lang F, Meyer TF. Acidic sphingomyelinase mediates entry of N. gonorrhoeae into nonphagocytic cells. Cell. 1997;91:605–15.
Grassme H, Riehle A, Wilker B, Gulbins E. Rhinoviruses infect human epithelial cells via ceramide-enriched membrane platforms. J Biol Chem. 2005;280:26256–62.
Grassmé H, Schwarz H, Gulbins E. Molecular mechanisms of ceramide-mediated CD95 clustering. Biochem Biophys Res Commun. 2001;284:1016–30.
Greve JM, Davis G, Meyer AM, Forte CP, Yost SC, Marlor CW, Kamarck ME, McClelland A. The major human rhinovirus receptor is ICAM-1. Cell. 1989;56:839–47.
Grignon N. Using SIMS and MIMS in biological materials: application to higher plants. Methods Mol Biol. 2007;369:569–91.
Grover R, Fischer J, Schwarz FW, Walter WJ, Schwille P, Diez S. Transport efficiency of membrane-anchored kinesin-1 motors depends on motor density and diffusivity. Proc Natl Acad Sci U S A. 2016;113:E7185–93.
Gruenberg J, van der Goot FG. Mechanisms of pathogen entry through the endosomal compartments. Nat Rev Mol Cell Biol. 2006;7:495–504.
Gulbins E, Dreschers S, Wilker B, Grassme H. Ceramide, membrane rafts and infections. J Mol Med (Berl). 2004;82:357–63.
Guo Q, Shen N, Yuan K, Li J, Wu H, Zeng Y, Fox 3rd J, Bansal AK, Singh BB, Gao H, Wu M. Caveolin-1 plays a critical role in host immunity against Klebsiella pneumoniae by regulating STAT5 and Akt activity. Eur J Immunol. 2012;42:1500–11.
Gutiérrez M, Isa P, Sánchez-San Martin C, Pérez-Vargas J, Espinosa R, Arias CF, López S. Different rotavirus strains enter MA104 cells through different endocytic pathways: the role of clathrin-mediated endocytosis. J Virol. 2010;84:9161–9.
Ha T. Single-molecule fluorescence methods for the study of nucleic acids. Curr Opin Struct Biol. 2001a;11:287–92.
Ha T. Single-molecule fluorescence resonance energy transfer. Methods. 2001b;25:78–86.
Hammache D, Pieroni G, Yahi N, Delezay O, Koch N, Lafont H, Tamalet C, Fantini J. Specific interaction of HIV-1 and HIV-2 surface envelope glycoproteins with monolayers of galactosylceramide and ganglioside GM3. J Biol Chem. 1998;273:7967–71.
Hannun YA, Obeid LM. The ceramide-centric universe of lipid-mediated cell regulation: stress encounters of the lipid kind. J Biol Chem. 2002;277:25847–50.
Hansen CG, Nichols BJ. Molecular mechanisms of clathrin-independent endocytosis. J Cell Sci. 2009;122:1713–21.
Harder T, Scheiffele P, Verkade P, Simons K. Lipid domain structure of the plasma membrane revealed by patching of membrane components. J Cell Biol. 1998;141:929–42.
Hartlova A, Cerveny L, Hubalek M, Krocova Z, Stulik J. Membrane rafts: a potential gateway for bacterial entry into host cells. Microbiol Immunol. 2010;54:237–45.
Hashimoto M, Hossain S, Yamasaki H, Yazawa K, Masumura S. Effects of eicosapentaenoic acid and docosahexaenoic acid on plasma membrane fluidity of aortic endothelial cells. Lipids. 1999;34:1297–304.
Hauck CR, Grassmé H, Bock J, Jendrossek V, Ferlinz K, Meyer TF, Gulbins E. Acid sphingomyelinase is involved in CEACAM receptor-mediated phagocytosis of Neisseria gonorrhoeae. FEBS Lett. 2000;478:260–6.
Hess ST, Girirajan TP, Mason MD. Ultra-high resolution imaging by fluorescence photoactivation localization microscopy. Biophys J. 2006;91:4258–72.
Heung LJ, Luberto C, Del Poeta M. Role of sphingolipids in microbial pathogenesis. Infect Immun. 2006;74:28–39.
Hill CM, Littman D. Identification of a major co-receptor for primary isolates of HIV-1. Nature. 1996;382:668–9.
Hinner MJ, Johnsson K. How to obtain labeled proteins and what to do with them. Curr Opin Biotechnol. 2010;21:766–76.
Holyoak CD, Stratford M, McMullin Z, Cole MB, Crimmins K, Brown AJ, Coote PJ. Activity of the plasma membrane H(+)-ATPase and optimal glycolytic flux are required for rapid adaptation and growth of Saccharomyces cerevisiae in the presence of the weak-acid preservative sorbic acid. Appl Environ Microbiol. 1996;62:3158–64.
Huang B, Bates M, Zhuang X. Super-resolution fluorescence microscopy. Annu Rev Biochem. 2009a;78:993–1016.
Huang H-C, Chen C-C, Chang W-C, Tao M-H, Huang C. Entry of hepatitis B virus into immortalized human primary hepatocytes by clathrin-dependent endocytosis. J Virol. 2012;86:9443–53.
Huang J, Ren T, Guan H, Jiang Y, Cheng H. HTLV-1 Tax is a critical lipid raft modulator that hijacks IkappaB kinases to the microdomains for persistent activation of NF-kappaB. J Biol Chem. 2009b;284:6208–17.
Huang H, Weaver A, Wu E, Li Y, Gao H, Fan W, Wu M. Lipid-based signaling modulates DNA repair response and survival against Klebsiella pneumoniae infection in host cells and in mice. Am J Respir Cell Mol Biol. 2013;49:798–807.
Hug P, Lin HM, Korte T, Xiao X, Dimitrov DS, Wang JM, Puri A, Blumenthal R. Glycosphingolipids promote entry of a broad range of human immunodeficiency virus type 1 isolates into cell lines expressing CD4, CXCR4, and/or CCR5. J Virol. 2000;74:6377–85.
Irani VR, Maslow JN. Induction of murine macrophage TNF-alpha synthesis by Mycobacterium avium is modulated through complement-dependent interaction via complement receptors 3 and 4 in relation to M. avium glycopeptidolipid. FEMS Microbiol Lett. 2005;246:221–8.
Jermy A. Bacterial lipid rafts discovered. Nat Rev Microbiol. 2010;8:756.
Jones KS, Fugo K, Petrow-Sadowski C, Huang Y, Bertolette DC, Lisinski I, Cushman SW, Jacobson S, Ruscetti FW. Human T-cell leukemia virus type 1 (HTLV-1) and HTLV-2 use different receptor complexes to enter T cells. J Virol. 2006;80:8291–302.
Jong A, Wu CH, Prasadarao NV, Kwon-Chung KJ, Chang YC, Ouyang Y, Shackleford GM, Huang SH. Invasion of Cryptococcus neoformans into human brain microvascular endothelial cells requires protein kinase C-alpha activation. Cell Microbiol. 2008;10:1854–65.
Kapadia SB, Barth H, Baumert T, McKeating JA, Chisari FV. Initiation of hepatitis C virus infection is dependent on cholesterol and cooperativity between CD81 and scavenger receptor B type I. J Virol. 2007;81:374–83.
Kayal S, Charbit A. Listeriolysin O: a key protein of Listeria monocytogenes with multiple functions. FEMS Microbiol Rev. 2006;30:514–29.
Kim S, Watarai M, Makino S, Shirahata T. Membrane sorting during swimming internalization of Brucella is required for phagosome trafficking decisions. Microb Pathog. 2002;33:225–37.
Kong KF, Vuong C, Otto M. Staphylococcus quorum sensing in biofilm formation and infection. Int J Med Microbiol. 2006;296:133–9.
Korade Z, Kenworthy AK. Lipid rafts, cholesterol, and the brain. Neuropharmacology. 2008;55:1265–73.
Korlach J, Schwille P, Webb WW, Feigenson GW. Characterization of lipid bilayer phases by confocal microscopy and fluorescence correlation spectroscopy. Proc Natl Acad Sci U S A. 1999;96:8461–6.
Kurzchalia T. Anthrax toxin rafts into cells. J Cell Biol. 2003;160:295–6.
Koshizuka T, Kawaguchi, Y, Nozawa N, Mori I, Nishiyama Y. Herpes simplex virus protein UL11 but not UL51 is associated with lipid rafts. Virus Genes. 2007;35:571-75.
Kusumi A, Ike H, Nakada C, Murase K, Fujiwara T. Single-molecule tracking of membrane molecules: plasma membrane compartmentalization and dynamic assembly of raft-philic signaling molecules. Semin Immunol. 2005b;17:3–21.
Kusumi A, Nakada C, Ritchie K, Murase K, Suzuki K, Murakoshi H, Kasai RS, Kondo J, Fujiwara T. Paradigm shift of the plasma membrane concept from the two-dimensional continuum fluid to the partitioned fluid: high-speed single-molecule tracking of membrane molecules. Annu Rev Biophys Biomol Struct. 2005a;34:351–78.
Kusumi A, Tsunoyama TA, Hirosawa KM, Kasai RS, Fujiwara TK. Tracking single molecules at work in living cells. Nat Chem Biol. 2014;10:524–32.
Kwiatek JM, Owen DM, Abu-Siniyeh A, Yan P, Loew LM, Gaus K. Characterization of a new series of fluorescent probes for imaging membrane order. PLoS One. 2013;8:e52960.
Lacalle RA, Mira E, Gomez-Mouton C, Jimenez-Baranda S, Martinez AC, Manes S. Specific SHP-2 partitioning in raft domains triggers integrin-mediated signaling via rho activation. J Cell Biol. 2002;157:277–89.
Lafont F, van der Goot FG. Bacterial invasion via lipid rafts. Cell Microbiol. 2005;7:613–20.
Lafont F, Tran Van Nhieu G, Hanada K, Sansonetti P, van der Goot FG. Initial steps of Shigella infection depend on the cholesterol/sphingolipid raft-mediated CD44-IpaB interaction. EMBO J. 2002;21:4449–57.
Lang PA, Kempe DS, Tanneur V, Eisele K, Klarl BA, Myssina S, Jendrossek V, Ishii S, Shimizu T, Waidmann M, Hessler G, Huber SM, Lang F, Wieder T. Stimulation of erythrocyte ceramide formation by platelet-activating factor. J Cell Sci. 2005;118:1233–43.
Laniosz V, Holthusen KA, Meneses PI. Bovine papillomavirus type 1: from clathrin to caveolin. J Virol. 2008;82:6288–98.
LaRocca TJ, Pathak P, Chiantia S, Toledo A, Silvius JR, Benach JL, London E. Proving lipid rafts exist: membrane domains in the prokaryote Borrelia burgdorferi have the same properties as eukaryotic lipid rafts. PLoS Pathog. 2013;9:e1003353.
Lattif AA, Mukherjee PK, Chandra J, Roth MR, Welti R, Rouabhia M, Ghannoum MA. Lipidomics of Candida albicans biofilms reveals phase-dependent production of phospholipid molecular classes and role for lipid rafts in biofilm formation. Microbiology. 2011;157:3232–42.
Lee S-H, Chung Y-H, Cho N-H, Gwack Y, Feng P, Jung JU. Modulation of T-cell receptor signal transduction by herpesvirus signaling adaptor protein. Mol Cell Biol. 2004;24:5369–82.
Lee CJ, Liao CL, Lin YL. Flavivirus activates phosphatidylinositol 3-kinase signaling to block caspase-dependent apoptotic cell death at the early stage of virus infection. J Virol. 2005;79:8388–99.
Leung K, Kim JO, Ganesh L, Kabat J, Schwartz O, Nabel GJ. HIV-1 assembly: viral glycoproteins segregate quantally to lipid rafts that associate individually with HIV-1 capsids and virions. Cell Host Microbe. 2008a;3:285–92.
Leung K, Kim J-O, Ganesh L, Kabat J, Schwartz O, Nabel GJ. Quantal formation of lentiviruses in cells: segregation of viral glycoproteins to lipid rafts that associate individually with HIV-1 Capsids. Cell Host Microbe. 2008b;3:285.
Li G-M, Li Y-G, Yamate M, Li S-M, Ikuta K. Lipid rafts play an important role in the early stage of severe acute respiratory syndrome-coronavirus life cycle. Microbes Infect. 2007;9:96–102.
Liao Z, Cimakasky LM, Hampton R, Nguyen DH, Hildreth JE. Lipid rafts and HIV pathogenesis: host membrane cholesterol is required for infection by HIV type 1. AIDS Res Hum Retrovir. 2001a;17:1009–19.
Liao Z, Cimakasky LM, Hampton R, Nguyen DH, Hildreth JE. Lipid rafts and HIV pathogenesis: host membrane cholesterol is required for infection by HIV type 1. AIDS Res Hum Retrovir. 2001b;17:1009–19.
Liao Z, Graham DR, Hildreth JE. Lipid rafts and HIV pathogenesis: virion-associated cholesterol is required for fusion and infection of susceptible cells. AIDS Res Hum Retrovir. 2003;19:675–87.
Lingwood D, Simons K. Lipid rafts as a membrane-organizing principle. Science. 2010;327:46–50.
Liu CI, Liu GY, Song Y, Yin F, Hensler ME, Jeng WY, Nizet V, Wang AH, Oldfield E. A cholesterol biosynthesis inhibitor blocks Staphylococcus aureus virulence. Science. 2008;319:1391–4.
Long M, Huang SH, Wu CH, Shackleford GM, Jong A. Lipid raft/caveolae signaling is required for Cryptococcus neoformans invasion into human brain microvascular endothelial cells. J Biomed Sci. 2012;19:19.
Lopez D. Molecular composition of functional microdomains in bacterial membranes. Chem Phys Lipids. 2015;192:3–11.
Lopez D, Kolter R. Functional microdomains in bacterial membranes. Genes Dev. 2010;24:1893–902.
Low H, Mukhamedova N, Cui HL, McSharry BP, Avdic S, Hoang A, Ditiatkovski M, Liu Y, Fu Y, Meikle PJ, Blomberg M, Polyzos KA, Miller WE, Religa P, Bukrinsky M, Soderberg-Naucler C, Slobedman B, Sviridov D. Cytomegalovirus restructures lipid rafts via a US28/CDC42-mediated pathway, enhancing cholesterol efflux from host cells. Cell Rep. 2016;16:186–200.
Lu YE, Kielian M. Semliki forest virus budding: assay, mechanisms, and cholesterol requirement. J Virol. 2000;74:7708–19.
Luberto C, Martinez-Marino B, Taraskiewicz D, Bolanos B, Chitano P, Toffaletti DL, Cox GM, Perfect JR, Hannun YA, Balish E, Del Poeta M. Identification of App1 as a regulator of phagocytosis and virulence of Cryptococcus neoformans. J Clin Invest. 2003;112:1080–94.
Ludwig A, Otto GP, Riento K, Hams E, Fallon PG, Nichols BJ. Flotillin microdomains interact with the cortical cytoskeleton to control uropod formation and neutrophil recruitment. J Cell Biol. 2010;191:771–81.
Luo C, Wang K, Liu DQ, Li Y, Zhao QS. The functional roles of lipid rafts in T cell activation, immune diseases and HIV infection and prevention. Cell Mol Immunol. 2008;5:1–7.
Ma DW, Seo J, Davidson LA, Callaway ES, Fan YY, Lupton JR, Chapkin RS. n-3 PUFA alter caveolae lipid composition and resident protein localization in mouse colon. FASEB J. 2004;18:1040–2.
Maguy A, Hebert TE, Nattel S. Involvement of lipid rafts and caveolae in cardiac ion channel function. Cardiovasc Res. 2006;69:798–807.
Maldonado-García G, Chico-Ortiz M, Lopez-Marin L, Sánchez-García FJ. High-polarity Mycobacterium avium-derived lipids interact with murine macrophage lipid rafts. Scand J Immunol. 2004;60:463–70.
Manes S, Ana Lacalle R, Gomez-Mouton C, Martinez AC. From rafts to crafts: membrane asymmetry in moving cells. Trends Immunol. 2003b;24:320–6.
Manes S, del Real G, Martinez AC. Pathogens: raft hijackers. Nat Rev Immunol. 2003a;3:557–68.
Manié SN, Debreyne S, Vincent S, Gerlier D. Measles virus structural components are enriched into lipid raft microdomains: a potential cellular location for virus assembly. J Virol. 2000;74:305–11.
Marjomaki V, Pietiainen V, Matilainen H, Upla P, Ivaska J, Nissinen L, Reunanen H, Huttunen P, Hyypia T, Heino J. Internalization of echovirus 1 in caveolae. J Virol. 2002;76:1856–65.
Martin SW, Konopka JB. Lipid raft polarization contributes to hyphal growth in Candida albicans. Eukaryot Cell. 2004;3:675–84.
Matrosovich M, Suzuki T, Hirabayashi Y, Garten W, Webster RG, Klenk H-D. Gangliosides are not essential for influenza virus infection. Glycoconj J. 2006;23:107–13.
Maza PK, Straus AH, Toledo MS, Takahashi HK, Suzuki E. Interaction of epithelial cell membrane rafts with Paracoccidioides brasiliensis leads to fungal adhesion and Src-family kinase activation. Microbes Infect. 2008;10:540–7.
McCubrey JA, Steelman LS, Chappell WH, Abrams SL, Wong EW, Chang F, Lehmann B, Terrian DM, Milella M, Tafuri A, Stivala F, Libra M, Basecke J, Evangelisti C, Martelli AM, Franklin RA. Roles of the Raf/MEK/ERK pathway in cell growth, malignant transformation and drug resistance. Biochim Biophys Acta. 2007;1773:1263–84.
McDonald TP, Pitt AR, Brown G, Rixon HWM, Sugrue RJ. Evidence that the respiratory syncytial virus polymerase complex associates with lipid rafts in virus-infected cells: a proteomic analysis. Virology. 2004;330:147–57.
Megha, London E. Ceramide selectively displaces cholesterol from ordered lipid domains (rafts): implications for lipid raft structure and function. J Biol Chem. 2004;279:9997–10004.
Melkonian KA, Ostermeyer AG, Chen JZ, Roth MG, Brown DA. Role of lipid modifications in targeting proteins to detergent-resistant membrane rafts. Many raft proteins are acylated, while few are prenylated. J Biol Chem. 1999;274:3910–7.
Miller ME, Adhikary S, Kolokoltsov AA, Davey RA. Ebolavirus requires acid sphingomyelinase activity and plasma membrane sphingomyelin for infection. J Virol. 2012;86:7473–83.
Minetti C, Sotgia F, Bruno C, Scartezzini P, Broda P, Bado M, Masetti E, Mazzocco M, Egeo A, Donati MA, Volonte D, Galbiati F, Cordone G, Bricarelli FD, Lisanti MP, Zara F. Mutations in the caveolin-3 gene cause autosomal dominant limb-girdle muscular dystrophy. Nat Genet. 1998;18:365–8.
Mishra S, Joshi PG. Lipid raft heterogeneity: an enigma. J Neurochem. 2007;103(Suppl 1):135–42.
Mo H, Elson CE. Studies of the isoprenoid-mediated inhibition of mevalonate synthesis applied to cancer chemotherapy and chemoprevention. Exp Biol Med (Maywood). 2004;229:567–85.
Monteiro da Silva JL, Andreotti PF, Benard G, Soares CP, Miranda ET, Mendes-Giannini MJ. Epithelial cells treated with genistein inhibit adhesion and endocytosis of Paracoccidioides brasiliensis. Antonie Van Leeuwenhoek. 2007;92:129–35.
Muller G. Oral delivery of protein drugs: driver for personalized medicine. Curr Issues Mol Biol. 2011;13:13–24.
Murakoshi H, Iino R, Kobayashi T, Fujiwara T, Ohshima C, Yoshimura A, Kusumi A. Single-molecule imaging analysis of Ras activation in living cells. Proc Natl Acad Sci U S A. 2004;101:7317–22.
Nagao T, Qin C, Grosheva I, Maxfield FR, Pierini LM. Elevated cholesterol levels in the plasma membranes of macrophages inhibit migration by disrupting RhoA regulation. Arterioscler Thromb Vasc Biol. 2007;27:1596–602.
Nanbo A, Kawanishi E, Yoshida R, Yoshiyama H. Exosomes derived from Epstein-Barr virus-infected cells are internalized via caveola-dependent endocytosis and promote phenotypic modulation in target cells. J Virol. 2013;87:10334–47.
Naranatt PP, Akula SM, Zien CA, Krishnan HH, Chandran B. Kaposi's sarcoma-associated herpesvirus induces the phosphatidylinositol 3-kinase-PKC-ζ-MEK-ERK signaling pathway in target cells early during infection: implications for infectivity. J Virol. 2003;77:1524–39.
Nayak DP, Hui EK, Barman S. Assembly and budding of influenza virus. Virus Res. 2004;106:147–65.
Nemerow GR, Wolfert R, McNaughton M, Cooper NR. Identification and characterization of the Epstein-Barr virus receptor on human B lymphocytes and its relationship to the C3d complement receptor (CR2). J Virol. 1985;55:347–51.
Ng CG, Coppens I, Govindarajan D, Pisciotta J, Shulaev V, Griffin DE. Effect of host cell lipid metabolism on alphavirus replication, virion morphogenesis, and infectivity. Proc Natl Acad Sci. 2008;105:16326–31.
Nguyen DH, Hildreth JE. Evidence for budding of human immunodeficiency virus type 1 selectively from glycolipid-enriched membrane lipid rafts. J Virol. 2000;74:3264–72.
Nguyen DH, Taub D. CXCR4 function requires membrane cholesterol: implications for HIV infection. J Immunol. 2002a;168:4121–6.
Nguyen DH, Taub D. CXCR4 function requires membrane cholesterol: implications for HIV infection. J Immunol. 2002b;168:4121–6.
Nicolson GL. The fluid-mosaic model of membrane structure: still relevant to understanding the structure, function and dynamics of biological membranes after more than 40 years. Biochim Biophys Acta. 2014;1838:1451–66.
Noisakran S, Dechtawewat T, Avirutnan P, Kinoshita T, Siripanyaphinyo U, Puttikhunt C, Kasinrerk W, Malasit P, Sittisombut N. Association of dengue virus NS1 protein with lipid rafts. J Gen Virol. 2008;89:2492–500.
Norkin LC, Anderson HA, Wolfrom SA, Oppenheim A. Caveolar endocytosis of simian virus 40 is followed by brefeldin A-sensitive transport to the endoplasmic reticulum, where the virus disassembles. J Virol. 2002a;76:5156–66.
Norkin LC, Anderson HA, Wolfrom SA, Oppenheim A. Caveolar endocytosis of simian virus 40 is followed by brefeldin A-sensitive transport to the endoplasmic reticulum, where the virus disassembles. J Virol. 2002b;76:5156–66.
Norkin LC, Wolfrom SA, Stuart ES. Association of caveolin with Chlamydia trachomatis inclusions at early and late stages of infection. Exp Cell Res. 2001;266:229–38.
Ohka S, Matsuda N, Tohyama K, Oda T, Morikawa M, Kuge S, Nomoto A. Receptor (CD155)-dependent endocytosis of poliovirus and retrograde axonal transport of the endosome. J Virol. 2004;78:7186–98.
Oka N, Yamamoto M, Schwencke C, Kawabe J, Ebina T, Ohno S, Couet J, Lisanti MP, Ishikawa Y. Caveolin interaction with protein kinase C. Isoenzyme-dependent regulation of kinase activity by the caveolin scaffolding domain peptide. J Biol Chem. 1997;272:33416–21.
Oliferenko S, Paiha K, Harder T, Gerke V, Schwarzler C, Schwarz H, Beug H, Gunthert U, Huber LA. Analysis of CD44-containing lipid rafts: recruitment of annexin II and stabilization by the actin cytoskeleton. J Cell Biol. 1999;146:843–54.
Ono A. Viruses and lipids. Viruses. 2010;2:1236–8.
Ono A, Freed EO. Role of lipid rafts in virus replication. Adv Virus Res. 2005;64:311–58.
Ono A, Waheed AA, Freed EO. Depletion of cellular cholesterol inhibits membrane binding and higher-order multimerization of human immunodeficiency virus type 1 gag. Virology. 2007;360:27–35.
Ono A, Waheed AA, Joshi A, Freed EO. Association of human immunodeficiency virus type 1 gag with membrane does not require highly basic sequences in the nucleocapsid: use of a novel Gag multimerization assay. J Virol. 2005;79:14131–40.
Ostrom RS, Liu X. Detergent and detergent-free methods to define lipid rafts and caveolae. Methods Mol Biol. 2007;400:459–68.
Ostrowski MA, Justement SJ, Catanzaro A, Hallahan CA, Ehler LA, Mizell SB, Kumar PN, Mican JA, Chun T-W, Fauci AS. Expression of chemokine receptors CXCR4 and CCR5 in HIV-1-infected and uninfected individuals. J Immunol. 1998;161:3195–201.
Palma-Guerrero J, Lopez-Jimenez JA, Perez-Berna AJ, Huang IC, Jansson HB, Salinas J, Villalain J, Read ND, Lopez-Llorca LV. Membrane fluidity determines sensitivity of filamentous fungi to chitosan. Mol Microbiol. 2010;75:1021–32.
Parihar SP, Guler R, Lang DM, Suzuki H, Marais AD, Brombacher F. Simvastatin enhances protection against Listeria monocytogenes infection in mice by counteracting listeria-induced phagosomal escape. PLoS One. 2013;8:e75490.
Partlow KC, Lanza GM, Wickline SA. Exploiting lipid raft transport with membrane targeted nanoparticles: a strategy for cytosolic drug delivery. Biomaterials. 2008;29:3367–75.
Parton RG, del Pozo MA. Caveolae as plasma membrane sensors, protectors and organizers. Nat Rev Mol Cell Biol. 2013;14:98–112.
Pasrija R, Panwar SL, Prasad R. Multidrug transporters CaCdr1p and CaMdr1p of Candida albicans display different lipid specificities: both ergosterol and sphingolipids are essential for targeting of CaCdr1p to membrane rafts. Antimicrob Agents Chemother. 2008;52:694–704.
Patel KP, Coyne CB, Bergelson JM. Dynamin-and lipid raft-dependent entry of decay-accelerating factor (DAF)-binding and non-DAF-binding coxsackieviruses into nonpolarized cells. J Virol. 2009;83:11064–77.
Peiffer I, Servin AL, Bernet-Camard MF. Piracy of decay-accelerating factor (CD55) signal transduction by the diffusely adhering strain Escherichia coli C1845 promotes cytoskeletal F-actin rearrangements in cultured human intestinal INT407 cells. Infect Immun. 1998;66:4036–42.
Pelkmans L. Secrets of caveolae- and lipid raft-mediated endocytosis revealed by mammalian viruses. Biochim Biophys Acta. 2005;1746:295–304.
Pelkmans L, Burli T, Zerial M, Helenius A. Caveolin-stabilized membrane domains as multifunctional transport and sorting devices in endocytic membrane traffic. Cell. 2004;118:767–80.
Pelkmans L, Kartenbeck J, Helenius A. Caveolar endocytosis of simian virus 40 reveals a new two-step vesicular-transport pathway to the ER. Nat Cell Biol. 2001;3:473–83.
Peters PJ, Mironov Jr A, Peretz D, van Donselaar E, Leclerc E, Erpel S, DeArmond SJ, Burton DR, Williamson RA, Vey M, Prusiner SB. Trafficking of prion proteins through a caveolae-mediated endosomal pathway. J Cell Biol. 2003;162:703–17.
Pierini LM, Eddy RJ, Fuortes M, Seveau S, Casulo C, Maxfield FR. Membrane lipid organization is critical for human neutrophil polarization. J Biol Chem. 2003;278:10831–41.
Pietiainen VM, Marjomaki V, Heino J, Hyypia T. Viral entry, lipid rafts and caveosomes. Ann Med. 2005;37:394–403.
Pike LJ. Lipid rafts: bringing order to chaos. J Lipid Res. 2003;44:655–67.
Pike LJ. Rafts defined: a report on the Keystone Symposium on Lipid Rafts and Cell Function. J Lipid Res. 2006;47:1597–8.
Pike LJ. The challenge of lipid rafts. J Lipid Res. 2009;50(Suppl):S323–8.
Pique C, Pham D, Tursz T, Dokhélar M-C. The cytoplasmic domain of the human T-cell leukemia virus type I envelope can modulate envelope functions in a cell type-dependent manner. J Virol. 1993;67:557–61.
Popik W, Alce TM, Au W-C. Human immunodeficiency virus type 1 uses lipid raft-colocalized CD4 and chemokine receptors for productive entry into CD4+ T cells. J Virol. 2002;76:4709–22.
Puerta-Guardo H, Mosso C, Medina F, Liprandi F, Ludert JE, del Angel RM. Antibody-dependent enhancement of dengue virus infection in U937 cells requires cholesterol-rich membrane microdomains. J Gen Virol. 2010;91:394–403.
Quirin K, Eschli B, Scheu I, Poort L, Kartenbeck J, Helenius A. Lymphocytic choriomeningitis virus uses a novel endocytic pathway for infectious entry via late endosomes. Virology. 2008;378:21–33.
Raghu H, Sharma-Walia N, Veettil MV, Sadagopan S, Caballero A, Sivakumar R, Varga L, Bottero V, Chandran B. Lipid rafts of primary endothelial cells are essential for Kaposi's sarcoma-associated herpesvirus/human herpesvirus 8-induced phosphatidylinositol 3-kinase and RhoA-GTPases critical for microtubule dynamics and nuclear delivery of viral DNA but dispensable for binding and entry. J Virol. 2007;81:7941–59.
Rai A, Pathak D, Thakur S, Singh S, Dubey AK, Mallik R. Dynein clusters into lipid microdomains on phagosomes to drive rapid transport toward lysosomes. Cell. 2016;164:722–34.
Rajendran L, Schneider A, Schlechtingen G, Weidlich S, Ries J, Braxmeier T, Schwille P, Schulz JB, Schroeder C, Simons M, Jennings G, Knolker HJ, Simons K. Efficient inhibition of the Alzheimer's disease beta-secretase by membrane targeting. Science. 2008;320:520–3.
Raulin J. Human immunodeficiency virus and host cell lipids. Interesting pathways in research for a new HIV therapy. Prog Lipid Res. 2002;41:27–65.
Razani B, Engelman JA, Wang XB, Schubert W, Zhang XL, Marks CB, Macaluso F, Russell RG, Li M, Pestell RG, Di Vizio D, Hou Jr H, Kneitz B, Lagaud G, Christ GJ, Edelmann W, Lisanti MP. Caveolin-1 null mice are viable but show evidence of hyperproliferative and vascular abnormalities. J Biol Chem. 2001;276:38121–38.
Reyes-del Valle J, Chávez-Salinas S, Medina F, del Angel RM. Heat shock protein 90 and heat shock protein 70 are components of dengue virus receptor complex in human cells. J Virol. 2005;79:4557–67.
Riethmuller J, Riehle A, Grassme H, Gulbins E. Membrane rafts in host-pathogen interactions. Biochim Biophys Acta. 2006;1758:2139–47.
Rollason R, Korolchuk V, Hamilton C, Schu P, Banting G. Clathrin-mediated endocytosis of a lipid-raft-associated protein is mediated through a dual tyrosine motif. J Cell Sci. 2007;120:3850–8.
Ronnebaumer K, Gross U, Bohne W. The nascent parasitophorous vacuole membrane of Encephalitozoon cuniculi is formed by host cell lipids and contains pores which allow nutrient uptake. Eukaryot Cell. 2008;7:1001–8.
Rossy J, Schlicht D, Engelhardt B, Niggli V. Flotillins interact with PSGL-1 in neutrophils and, upon stimulation, rapidly organize into membrane domains subsequently accumulating in the uropod. PLoS One. 2009;4:e5403.
Rothberg KG, Heuser JE, Donzell WC, Ying YS, Glenney JR, Anderson RG. Caveolin, a protein component of caveolae membrane coats. Cell. 1992;68:673–82.
Roux PP, Blenis J. ERK and p38 MAPK-activated protein kinases: a family of protein kinases with diverse biological functions. Microbiol Mol Biol Rev. 2004;68:320–44.
Rust MJ, Bates M, Zhuang X. Sub-diffraction-limit imaging by stochastic optical reconstruction microscopy (STORM). Nat Methods. 2006;3:793–5.
Sahl SJ, Moerner WE. Super-resolution fluorescence imaging with single molecules. Curr Opin Struct Biol. 2013;23:778–87.
Sandvig K, van Deurs B. Transport of protein toxins into cells: pathways used by ricin, cholera toxin and Shiga toxin. FEBS Lett. 2002a;529:49–53.
Sandvig K, van Deurs B. Membrane traffic exploited by protein toxins. Annu Rev Cell Dev Biol. 2002b;18:1–24.
Sapin C, Colard O, Delmas O, Tessier C, Breton M, Enouf V, Chwetzoff S, Ouanich J, Cohen J, Wolf C, Trugnan G. Rafts promote assembly and atypical targeting of a nonenveloped virus, rotavirus, in Caco-2 cells. J Virol. 2002;76:4591–602.
Schley PD, Brindley DN, Field CJ. (n-3) PUFA alter raft lipid composition and decrease epidermal growth factor receptor levels in lipid rafts of human breast cancer cells. J Nutr. 2007;137:548–53.
Schutz GJ, Kada G, Pastushenko VP, Schindler H. Properties of lipid microdomains in a muscle cell membrane visualized by single molecule microscopy. EMBO J. 2000;19:892–901.
Selvarangan R, Goluszko P, Popov V, Singhal J, Pham T, Lublin DM, Nowicki S, Nowicki B. Role of decay-accelerating factor domains and anchorage in internalization of Dr-fimbriated Escherichia coli. Infect Immun. 2000;68:1391–9.
Sengupta P, Jovanovic-Talisman T, Skoko D, Renz M, Veatch SL, Lippincott-Schwartz J. Probing protein heterogeneity in the plasma membrane using PALM and pair correlation analysis. Nat Methods. 2011;8:969–75.
Seveau S, Bierne H, Giroux S, Prevost MC, Cossart P. Role of lipid rafts in E-cadherin-- and HGF-R/met--mediated entry of Listeria monocytogenes into host cells. J Cell Biol. 2004;166:743–53.
Sharp EL, Davis-Poynter NJ, Farrell HE. Analysis of the subcellular trafficking properties of murine cytomegalovirus M78, a 7 transmembrane receptor homologue. J Gen Virol. 2009;90:59–68.
Shukla D, Spear PG. Herpesviruses and heparan sulfate: an intimate relationship in aid of viral entry. J Clin Invest. 2001;108:503–10.
Siafakas AR, Wright LC, Sorrell TC, Djordjevic JT. Lipid rafts in Cryptococcus neoformans concentrate the virulence determinants phospholipase B1 and Cu/Zn superoxide dismutase. Eukaryot Cell. 2006;5:488–98.
Silva LC, Futerman AH, Prieto M. Lipid raft composition modulates sphingomyelinase activity and ceramide-induced membrane physical alterations. Biophys J. 2009;96:3210–22.
Silva BM, Sousa LP, Gomes-Ruiz AC, Leite FG, Teixeira MM, da Fonseca FG, Pimenta PF, Ferreira PC, Kroon EG, Bonjardim CA. The dengue virus nonstructural protein 1 (NS1) increases NF-kappaB transcriptional activity in HepG2 cells. Arch Virol. 2011;156:1275–9.
Silvius JR, Nabi IR. Fluorescence-quenching and resonance energy transfer studies of lipid microdomains in model and biological membranes. Mol Membr Biol. 2006;23:5–16.
Simonetti N, Strippoli V, Cassone A. Yeast-mycelial conversion induced by N-acetyl-D-glucosamine in Candida albicans. Nature. 1974;250:344–6.
Simons K, Gruenberg J. Jamming the endosomal system: lipid rafts and lysosomal storage diseases. Trends Cell Biol. 2000;10:459–62.
Simons K, Ikonen E. Functional rafts in cell membranes. Nature. 1997;387:569–72.
Simons K, Toomre D. Lipid rafts and signal transduction. Nat Rev Mol Cell Biol. 2000;1:31–9.
Singer SJ, Nicolson GL. The fluid mosaic model of the structure of cell membranes. Science. 1972;175:720–31.
Sinha B, François PP, Nüße O, Foti M, Hartford OM, Vaudaux P, Foster TJ, Lew DP, Herrmann M, Krause KH. Fibronectin-binding protein acts as Staphylococcus aureus invasin via fibronectin bridging to integrin α5β1. Cell Microbiol. 1999;1:101–17.
Siskind LJ, Kolesnick RN, Colombini M. Ceramide channels increase the permeability of the mitochondrial outer membrane to small proteins. J Biol Chem. 2002;277:26796–803.
Somani VK, Aggarwal S, Singh D, Prasad T, Bhatnagar R. Identification of novel raft marker protein, FlotP in Bacillus anthracis. Front Microbiol. 2016;7:169.
Sowa G, Xie L, Xu L, Sessa WC. Serine 23 and 36 phosphorylation of caveolin-2 is differentially regulated by targeting to lipid raft/caveolae and in mitotic endothelial cells. Biochemistry. 2008;47:101–11.
Spiegel S, Milstien S. Sphingosine-1-phosphate: an enigmatic signalling lipid. Nat Rev Mol Cell Biol. 2003;4:397–407.
Stang E, Kartenbeck J, Parton RG. Major histocompatibility complex class I molecules mediate association of SV40 with caveolae. Mol Biol Cell. 1997;8:47–57.
Staunton DE, Merluzzi VJ, Rothlein R, Barton R, Marlin SD, Springer TA. A cell adhesion molecule, ICAM-1, is the major surface receptor for rhinoviruses. Cell. 1989;56:849–53.
Steinberg BE, Grinstein S. Pathogen destruction versus intracellular survival: the role of lipids as phagosomal fate determinants. J Clin Invest. 2008;118:2002–11.
Stuart AD, Eustace HE, McKee TA, Brown T. A novel cell entry pathway for a DAF-using human enterovirus is dependent on lipid rafts. J Virol. 2002a;76:9307–22.
Stuart AD, Eustace HE, McKee TA, Brown TD. A novel cell entry pathway for a DAF-using human enterovirus is dependent on lipid rafts. J Virol. 2002b;76:9307–22.
Suzuki T, Suzuki Y. Virus infection and lipid rafts. Biol Pharm Bull. 2006;29:1538–41.
Suzuki Y, Ito T, Suzuki T, Holland RE, Chambers TM, Kiso M, Ishida H, Kawaoka Y. Sialic acid species as a determinant of the host range of influenza A viruses. J Virol. 2000;74:11825–31.
Swamy MJ, Ciani L, Ge M, Smith AK, Holowka D, Baird B, Freed JH. Coexisting domains in the plasma membranes of live cells characterized by spin-label ESR spectroscopy. Biophys J. 2006;90:4452–65.
Tagliari L, Toledo MS, Lacerda TG, Suzuki E, Straus AH, Takahashi HK. Membrane microdomain components of Histoplasma capsulatum yeast forms, and their role in alveolar macrophage infectivity. Biochim Biophys Acta. 2012;1818:458–66.
Takahashi T, Suzuki T. Function of membrane rafts in viral lifecycles and host cellular response. Biochem Res Int. 2011a;2011:245090.
Takahashi T, Suzuki T. Function of membrane rafts in viral lifecycles and host cellular response. Biochem Res Int. 2011b;2011.
Tang H, Kawabata A, Takemoto M, Yamanishi K, Mori Y. Human herpesvirus-6 infection induces the reorganization of membrane microdomains in target cells, which are required for virus entry. Virology. 2008;378:265–71.
Tateyama M, Aoki M, Nishino I, Hayashi YK, Sekiguchi S, Shiga Y, Takahashi T, Onodera Y, Haginoya K, Kobayashi K, Iinuma K, Nonaka I, Arahata K, Itoyama Y. Mutation in the caveolin-3 gene causes a peculiar form of distal myopathy. Neurology. 2002;58:323–5.
Tikhomirov O, Carpenter G. Ligand-induced, p38-dependent apoptosis in cells expressing high levels of epidermal growth factor receptor and ErbB-2. J Biol Chem. 2004;279:12988–96.
Triantafilou K, Triantafilou M. Lipid raft microdomains: key sites for coxsackievirus A9 infectious cycle. Virology. 2003;317:128–35.
Triantafilou K, Triantafilou M. Lipid-raft-dependent coxsackievirus B4 internalization and rapid targeting to the Golgi. Virology. 2004;326:6–19.
Triantafilou K, Fradelizi D, Wilson K, Triantafilou M. GRP78, a coreceptor for coxsackievirus A9, interacts with major histocompatibility complex class I molecules which mediate virus internalization. J Virol. 2002;76:633–43.
Tsai B, Gilbert JM, Stehle T, Lencer W, Benjamin TL, Rapoport TA. Gangliosides are receptors for murine polyoma virus and SV40. EMBO J. 2003a;22:4346–55.
Tsai B, Gilbert JM, Stehle T, Lencer W, Benjamin TL, Rapoport TA. Gangliosides are receptors for murine polyoma virus and SV40. EMBO J. 2003b;22:4346–55.
Turk HF, Chapkin RS. Membrane lipid raft organization is uniquely modified by n-3 polyunsaturated fatty acids. Prostaglandins Leukot Essent Fatty Acids. 2013;88:43–7.
Upla P, Marjomaki V, Kankaanpaa P, Ivaska J, Hyypia T, Van Der Goot FG, Heino J. Clustering induces a lateral redistribution of alpha 2 beta 1 integrin from membrane rafts to caveolae and subsequent protein kinase C-dependent internalization. Mol Biol Cell. 2004;15:625–36.
van Meer G, Poorthuis BJ, Wirtz KW, Op den Kamp JA, van Deenen LL. Transbilayer distribution and mobility of phosphatidylcholine in intact erythrocyte membranes. A study with phosphatidylcholine exchange protein. Eur J Biochem. 1980;103:283–8.
van Meer G, Stelzer EH, Wijnaendts-van-Resandt RW, Simons K. Sorting of sphingolipids in epithelial (Madin-Darby canine kidney) cells. J Cell Biol. 1987;105:1623–35.
van Meer G, Voelker DR, Feigenson GW. Membrane lipids: where they are and how they behave. Nat Rev Mol Cell Biol. 2008;9:112–24.
Vaughan CJ. Prevention of stroke and dementia with statins: effects beyond lipid lowering. Am J Cardiol. 2003;91:23B–9B.
Vennema H, Godeke GJ, Rossen JW, Voorhout WF, Horzinek MC, Opstelten DJ, Rottier PJ. Nucleocapsid-independent assembly of coronavirus-like particles by co-expression of viral envelope protein genes. EMBO J. 1996;15:2020–8.
Vieira FS, Correa G, Einicker-Lamas M, Coutinho-Silva R. Host-cell lipid rafts: a safe door for micro-organisms? Biol Cell. 2010;102:391–407.
Wang H. Lipid rafts: a signaling platform linking cholesterol metabolism to synaptic deficits in autism spectrum disorders. Front Behav Neurosci. 2014;8:104.
Wang M, Hajishengallis G. Lipid raft-dependent uptake, signalling and intracellular fate of Porphyromonas gingivalis in mouse macrophages. Cell Microbiol. 2008;10:2029–42.
Wang L, Jia Y, Tang RJ, Xu Z, Cao YB, Jia XM, Jiang YY. Proteomic analysis of Rta2p-dependent raft-association of detergent-resistant membranes in Candida albicans. PLoS One. 2012;7:e37768.
Wang H, Yang P, Liu K, Guo F, Zhang Y, Zhang G, Jiang C. SARS coronavirus entry into host cells through a novel clathrin-and caveolae-independent endocytic pathway. Cell Res. 2008;18:290–301.
Wang XM, Zhang Y, Kim HP, Zhou Z, Feghali-Bostwick CA, Liu F, Ifedigbo E, Xu X, Oury TD, Kaminski N, Choi AM. Caveolin-1: a critical regulator of lung fibrosis in idiopathic pulmonary fibrosis. J Exp Med. 2006;203:2895–906.
Watarai M, Derre I, Kirby J, Growney JD, Dietrich WF, Isberg RR. Legionella pneumophila is internalized by a macropinocytotic uptake pathway controlled by the Dot/Icm system and the mouse Lgn1 locus. J Exp Med. 2001;194:1081–96.
Whitt MA, Buonocore L, Prehaud C, Rose JK. Membrane fusion activity, oligomerization, and assembly of the rabies virus glycoprotein. Virology. 1991;185:681–8.
Wu B, Geng S, Bi Y, Liu H, Hu Y, Li X, Zhang Y, Zhou X, Zheng G, He B, Wang B. Herpes simplex virus 1 suppresses the function of lung dendritic cells via caveolin-1. Clin Vaccine Immunol. 2015;22:883–95.
Xiao F, McKeating JA, Baumert TF. A bile acid transporter as a candidate receptor for hepatitis B and D virus entry. J Hepatol. 2013;58:1246–8.
Xing L, Huhtala M, Pietiainen V, Kapyla J, Vuorinen K, Marjomaki V, Heino J, Johnson MS, Hyypia T, Cheng RH. Structural and functional analysis of integrin alpha2I domain interaction with echovirus 1. J Biol Chem. 2004;279:11632–8.
Yamamoto M, Toya Y, Schwencke C, Lisanti MP, Myers Jr MG, Ishikawa Y. Caveolin is an activator of insulin receptor signaling. J Biol Chem. 1998;273:26962–8.
Yao Y, Hong S, Zhou H, Yuan T, Zeng R, Liao K. The differential protein and lipid compositions of noncaveolar lipid microdomains and caveolae. Cell Res. 2009;19:497–506.
Yavlovich A, Kohen R, Ginsburg I, Rottem S. The reducing antioxidant capacity of Mycoplasma fermentans. FEMS Microbiol Lett. 2006;259:195–200.
Yoon C-S, Kim K-D, Park S-N, Cheong S-W. α 6 integrin is the main receptor of human papillomavirus type 16 VLP. Biochem Biophys Res Commun. 2001;283:668–73.
Yu C, Alterman M, Dobrowsky RT. Ceramide displaces cholesterol from lipid rafts and decreases the association of the cholesterol binding protein caveolin-1. J Lipid Res. 2005;46:1678–91.
Yu J, Fischman DA, Steck TL. Selective solubilization of proteins and phospholipids from red blood cell membranes by nonionic detergents. J Supramol Struct. 1973;1:233–48.
Zaas DW, Duncan M, Rae Wright J, Abraham SN. The role of lipid rafts in the pathogenesis of bacterial infections. Biochim Biophys Acta. 2005;1746:305–13.
Zhang HM, Li Z, Tsudome M, Ito S, Takami H, Horikoshi K. An alkali-inducible flotillin-like protein from Bacillus halodurans C-125. Protein J. 2005;24:125–31.
Zhang J, Pekosz A, Lamb RA. Influenza virus assembly and lipid raft microdomains: a role for the cytoplasmic tails of the spike glycoproteins. J Virol. 2000;74:4634–44.
Zhang J, Randall G, Higginbottom A, Monk P, Rice CM, McKeating JA. CD81 is required for hepatitis C virus glycoprotein-mediated viral infection. J Virol. 2004;78:1448–55.
Zhao F, Zhang J, Liu YS, Li L, He YL. Research advances on flotillins. Virol J. 2011;8:479.
Zheng W, Kollmeyer J, Symolon H, Momin A, Munter E, Wang E, Kelly S, Allegood JC, Liu Y, Peng Q, Ramaraju H, Sullards MC, Cabot M, Merrill Jr AH. Ceramides and other bioactive sphingolipid backbones in health and disease: lipidomic analysis, metabolism and roles in membrane structure, dynamics, signaling and autophagy. Biochim Biophys Acta. 2006;1758:1864–84.
Zhong Y, Smart EJ, Weksler B, Couraud PO, Hennig B, Toborek M. Caveolin-1 regulates human immunodeficiency virus-1 Tat-induced alterations of tight junction protein expression via modulation of the Ras signaling. J Neurosci. 2008;28:7788–96.
Zhu Y-Z, Cao M-M, Wang W-B, Wang W, Ren H, Zhao P, Qi Z-T. Association of heat-shock protein 70 with lipid rafts is required for Japanese encephalitis virus infection in Huh7 cells. J Gen Virol. 2012;93:61–71.
Acknowledgements
This work was supported by Young Clinical Scientist Award from the Flight Attendant Medical Research Institute (FAMRI-123253_YCSA_Faculty), NIH R15 (7 R15 ES023151 02), Southern University Foundation Grant (FY2017-017), and LBRN Startup Grant (2P20GM103424-14-Subaward No. 100011) to SB.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Prathyusha Bagam and Dhirendra P. Singh contributed equally.
Rights and permissions
About this article

Cite this article
Bagam, P., Singh, D.P., Inda, M.E. et al. Unraveling the role of membrane microdomains during microbial infections. Cell Biol Toxicol 33, 429–455 (2017). https://doi.org/10.1007/s10565-017-9386-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10565-017-9386-9