
Protein Disorder in Plant Stress Adaptation: From Late Embryogenesis Abundant to Other Intrinsically Disordered Proteins

Department of Biochemistry and Metabolism, John Innes Centre, Norwich Research Park, Norwich NR4 7UH, UK
Int. J. Mol. Sci. 2024, 25(2), 1178; https://doi.org/10.3390/ijms25021178
Submission received: 29 October 2023
/
Revised: 15 January 2024
/
Accepted: 16 January 2024
/
Published: 18 January 2024
(This article belongs to the Special Issue Genes Function and Mechanism Identification in Plant Stress Resistance 2.0)
Abstract
:Global climate change has caused severe abiotic and biotic stresses, affecting plant growth and food security. The mechanical understanding of plant stress responses is critical for achieving sustainable agriculture. Intrinsically disordered proteins (IDPs) are a group of proteins without unique three-dimensional structures. The environmental sensitivity and structural flexibility of IDPs contribute to the growth and developmental plasticity for sessile plants to deal with environmental challenges. This article discusses the roles of various disordered proteins in plant stress tolerance and resistance, describes the current mechanistic insights into unstructured proteins such as the disorder-to-order transition for adopting secondary structures to interact with specific partners (i.e., cellular membranes, membrane proteins, metal ions, and DNA), and elucidates the roles of liquid–liquid phase separation driven by protein disorder in stress responses. By comparing IDP studies in animal systems, this article provides conceptual principles of plant protein disorder in stress adaptation, reveals the current research gaps, and advises on the future research direction. The highlighting of relevant unanswered questions in plant protein disorder research aims to encourage more studies on these emerging topics to understand the mechanisms of action behind their stress resistance phenotypes.
1. Introduction
Human activities have released large amounts of greenhouse gases into the atmosphere, causing climate change [1]. Global climate change is the main cause of abiotic and biotic stresses for plants that include flooding, drought, heat, cold, salinity, pests, and microbes [2]. These environmental stresses have greatly influenced plant physiological processes and have adverse effects on agriculture productivity [2,3]. To tackle the environmental stresses, sessile plants have evolved multiple processes including stress sensing, hormone regulation, signal transduction, gene expression, and regulatory pathways to adjust growth and development in a spatiotemporal manner [4,5]. Understanding the mechanism of plant stress adaptation to adverse environmental conditions will open new opportunities for agricultural applications and global food security.
Intrinsically disordered proteins (IDPs) are a group of proteins natively lacking defined three-dimensional structures. The peculiarities of their amino acid sequences are known to be depleted in order-promoting residues (tryptophan, cysteine, tyrosine, isoleucine, phenylalanine, valine, asparagine, and leucine) and enriched in disorder-promoting residues (arginine, proline, glutamine, glycine, glutamate, serine, alanine, and lysine) [6,7]. The amino acid biases contribute their exceptional spatiotemporal heterogeneity and low conformational stability, which drive their atypical response to changing environments [6,7]. IDPs can gain structures in the presence of various osmolytes, under the changes in temperature and pH, and perform a disorder-to-order transition when interacting with binding partners (i.e., cellular membranes, proteins, metal ions, and DNA) [6,7,8,9,10]. These features allow IDPs to sense environmental changes, mediate corresponding signalling pathways, and control and fine-tune plant metabolism in response to light, mechanical forces, pH, redox potential, and drought/salt concentration [11,12,13]. Thus, IDPs are thought to play critical roles in dynamic and plastic responses for the survival of sessile plants under the constantly changing environment [14,15,16].
The great importance of IDPs in stress adaptation is shown by extremophiles (i.e., anhydrobiotic tardigrades and resurrection plants) exhibiting extreme tolerance to various environmental stressors. Tardigrades survive in adverse environments via a set of highly disordered proteins that protect biomaterial through vitrification [17,18]. Two plants with relatively high protein disorder abundance, resurrection grass (Oropetium thomaeum) and switch grass (Panicum virgatum), are both stress-tolerant [19]. A proteomics analysis revealed that various IDPs were induced in the resurrection plant Haberlea rhodopensis during stress responses [20], whereas IDPs in wheat and barley are mainly involved in regulating cellular and biological processes in response to stress [21]. The function of IDPs in protecting biomolecules under stress conditions has been proposed [22], but the precise mechanisms of their action are still largely unknown.
Intrinsically disordered regions (IDRs) of transcription factors are highly adaptive and are proposed to provide functional versatility in molecular recognition via their binding plasticity [22,23,24]. IDRs can also serve as signalling hubs that regulate a diverse array of signal transduction pathways [9,25]. IDPs/IDRs are key triggers of liquid–liquid phase separation (LLPS) to form biomolecular condensates, which allow the spatiotemporal organization of biochemical reactions by concentrating macromolecules locally [26,27]. In animal cells, biomolecular condensates contribute to the biogenesis of macromolecular machineries essential for gene expression, the sequestration of specific factors to regulate cellular processes, and the interconnection of various diseases and innate immunity [28,29]. Recent studies revealed that plant biomolecular condensates driven by IDRs switch the defence programming through sequestration and regulate translation and protein quality control machineries in the plant immune response [30,31,32]. The study of LLPS is still an emerging area in plant research [33], and future research providing more mechanistic insights into how LLPS is involved in plant stress adaptation is expected.
This article summarizes the current knowledge of the physiological roles of plant IDPs in abiotic and biotic stress responses and discusses their potential molecular functions, highlighting the structure–function relationship of IDPs, interactions between IDPs and their partners, and the role of LLPS under various stress conditions. This article proposes the conceptual principles of how the structural flexibility of protein disorder leads to plant stress adaptation and pinpoints unanswered but relevant questions and the potential approaches to overcome the current limitation of plant IDP research.
2. Late Embryogenesis Abundant (LEA) Proteins Confer Abiotic Stress Tolerance
Plant seeds are desiccation-tolerant organs and physiologically similar to anhydrobiosis [34]. During the late stages of seed maturation, a group of IDPs known as LEA proteins are highly expressed in plant seeds before they enter the desiccation phase [35,36]. LEA proteins are thought to be involved in desiccation tolerance by modulating various clients [10,35,36,37]. LEA proteins are widely studied in model and crop plants such as rice, tomato, and Arabidopsis. There are 34, 60, and 51 LEA genes identified in rice [38], tomato [39], and Arabidopsis [40], respectively. LEA genes in plants are non-randomly distributed within the chromosomes, and segmental and tandem duplications drive LEA gene expansion in the genome during evolution [39,41,42].
Most LEA genes have elements responding to abscisic acid (ABA) and/or a low temperature in their promoter regions, such as ABA response elements (ABREs) and C-repeats (CRTs), which agrees well with LEA genes being induced by abiotic stresses such as drought, salinity, heat, and freezing in vegetative tissues [40]. Drought and salinity stresses trigger ABA signalling, whereas sucrose-nonfermenting-1-related protein kinases (SnRK2s) function upstream of the transcription factors ABA-INSENSITIVE 3 (ABI3), ABI5, and ABFs (ABA-responsive element-binding factors) to regulate the LEA gene expression via ABRE [43,44]. Cold stress activates calcium signalling and the mitogen-activated protein kinase cascade pathway, whereas CBFs (C-repeat binding factors) regulate the expression of the group 2 LEA dehydrins via CRT [44] (Table 1). Most LEA proteins are part of a more widespread group of proteins called “hydrophilins”, which are desiccation-related IDPs and have been discovered in archeal, eubacterial, and eukaryotic domains, including yeast, nematodes, tardigrades, and plants [34,45,46,47]. Hydrophilins are proposed to be stress effectors against desiccation in anhydrobiotes [48]; therefore, it is not surprising that LEA proteins are often involved in various stress responses such as desiccation resistance in plants [37,44,49].
There are several well-known examples of LEA proteins in stress responses. Arabidopsis dehydrins EARLY RESPONSE TO DEHYDRATION 10 (ERD10) and ERD14 function as chaperones to protect cells under a high salinity, drought, and low temperature [50]; LOW-TEMPERATURE-INDUCED 30 (Lti30) protects the membrane during cold and dehydration stress [51,52]; the group 3 LEA protein COLD-REGULATED 15A (COR15A) confers freezing tolerance by interacting with the membrane [53]; and legume physiologically mature (PM) proteins, which belong to various LEA groups, function in abiotic stress tolerance [54,55,56]. Their proposed mechanism is discussed in the following sections. Besides the reviews summarizing dehydrins in abiotic stress tolerance in various plant species [44,49], recent reports showed that heterologous expression of XsLEA1-8 from the monocot resurrection plant Xerophyta schlechteri [57], MsLEA-D34 from alfalfa (Medicago sativa L.) [58], and dehydrin CdDHN4 from bermudagrass (Cynodon dactylon × Cynodon transvaalensis) [59] increased osmotic and salt tolerance in Arabidopsis. These reports suggest that in general, stress-responsive LEA proteins confer stress tolerance among different plant species.
The analysis of LEA proteins in Arabidopsis, tomato, and orchid revealed their wide subcellular distribution [41,60,61]. Experimental data showed Arabidopsis LEA proteins localized to the cytosol, nucleus, mitochondria, plastid, ER, and pexophagosome [60]. Most of the tomato LEA proteins were predicted to target the nucleus or cytoplasm, with some in the mitochondria, chloroplasts, or extracellular matrix [41]. As well, subcellular localization prediction indicated that orchid LEA proteins are located in the nucleus, cytoplasm, chloroplasts, and mitochondria [61]. LEA proteins are proposed to protect the integrity of membranes and the activity of enzymes and biomolecules under stress conditions [22]. Therefore, their ubiquitous expression might provide protection to the corresponding membranes of various organelles as well as enzymes and sequestering targets localized in different cellular compartments under certain stress conditions [35,60]. Multiple cellular locations were also observed in another IDP, the Stress and Growth Interconnector (SGI), from the oil crop rapeseed (Brassica napus), which enhanced biomass and yield under drought conditions via multifaceted interactions with catalases and dehydrins [62]. A wheat IDP, the Triticum aestivum Fusarium Resistance Orphan Gene (TaFROG), showed spatial transition by changing its nucleus localization to cytosolic bodies when interacting with SnRK1 [63]. Thus, the spatial flexibility of IDPs implies their versatile functions in dual/multiple subcellular localizations.
Although more novel stress-responsive IDPs in plants are being investigated, LEA proteins are still the most-studied plant IDP family (Table 2). Despite overwhelming functional evidence suggesting that LEA proteins are widespread mediators of abiotic stress tolerance in plants, their mechanisms of protection are largely unknown. Speculation and preliminary studies suggested that LEA proteins could interact with vastly different target biomolecules during stress conditions, such as molecular shielding for enzyme protection, membrane stabilization via lipid binding, and interactions with various clients as described in the following sections.

{kind=link}
{kind=link}
{kind=link}
Table 1.
Hormone regulation of IDPs/IDRs mentioned in this article.
| Hormone | Regulation of IDP/IDR Function in Stress Responses | Stressor Types | References |
|---|---|---|---|
| ABA | The expression of many LEA genes in plants is closely regulated by ABA and the corresponding signalling pathways. The promoter regions of LEA genes bear ABREs and CRTs, which can be recognized by various transcription factors such as ABI3, ABI5, ABFs, and CBPs during drought, salinity, and cold stress. | Abiotic stress | [43,44] |
| ABA | Rice RePRP is induced by water deficit and ABA in the root elongation zone and is sufficient and necessary for repression of root development by water deficit or ABA. | Abiotic stress | [64] |
| ABA, BR | Rice REM4.1 is transcriptionally upregulated by ABA and inhibits the formation and activation of a BR receptor kinase (BRI1-SERK1) complex, serving as a link between the ABA and BR signalling pathways. | Abiotic stress | [65] |
| SA | The major defence hormone SA triggers AtREM1.2/1.3-dependent membrane lipid nanodomain assembly, leading to plasmodesmata closure to impede virus spreading. | Biotic stress | [66] |
| SA | SA rapidly triggers the formation of nuclear GBPL3 condensates, which reprogram gene expression for disease resistance. | Biotic stress | [67] |
| SA | SA induces NPR1 condensates in cytoplasm to sequester and degrade stress proteins involved in cell death, promoting cell survival during the immune response. | Biotic stress | [30] |
| SA | SA induces massive formation of HEM1 condensates to restrict the availability of translation components and prevent immune gene translation during ETI. | Biotic stress | [32] |
IDPs/IDRs, intrinsically disordered proteins/intrinsically disordered regions; ABA, abscisic acid; LEA, late embryogenesis abundant; ABREs, ABA response elements; CRTs, C-repeats; ABI3, ABA-INSENSITIVE 3; ABI5, ABA-INSENSITIVE 5; ABFs, ABA-responsive element-binding factors; CBPs, C-repeat binding factors; RePRP, repetitive pro-rich protein; BR, brassinosteroid; REM4.1, REMORIN 4.1; BRI1, brassinosteroid-insensitive 1; SERK1, somatic embryogenesis receptor-like kinase 1; SA, salicylic acid; GBPL3, guanylate-binding protein (GBP)-like GTPase 3; NPR1, NONEXPRESSOR-OF-PATHOGENESIS-RELATED GENE 1; HEM1, named for heme biosynthesis in the yeast homologue; ETI, effector-triggered immunity.
3. Molecular Shielding for Enzyme Protection
Many studies have shown that plant IDPs (mainly LEA proteins) are able to protect lactate dehydrogenase enzyme activity against damage caused by desiccation, heat stress, or freezing and thawing cycles [59,68,69,70,71,72,73,74,75,76]. The disorder and flexibility structure of LEA proteins may allow them to act as a kind of “molecular shield”, forming a physical barrier between neighbouring macromolecules and preventing their aggregation and inactivation under stress conditions [77]. LEA proteins can act sterically as noninteracting space fillers and/or interact with client proteins electrostatically via polar and charged amino acids. This molecular shield function is proposed to be distinct from classical molecular chaperones assisting protein folding via hydrophobic interactions and tight complex formation. However, both protect client targets against intramolecular damage [78]. Polyethylene glycols (PEGs) are similar to dehydrins, the group 2 LEA proteins, in terms of polar polymeric chains with no appreciable structure. Both dehydrins and PEGs showed size-dependent enzyme cryoprotection, which suggests that the disorder and polar nature along with the hydrodynamic radius of the dehydrin contribute to the molecular shield mechanism [79,80,81]. Group 5 LEA proteins are atypical because they contain a high content of hydrophobic residues and are proposed to be natively folded [45]. However, the group 5 LEA protein, Medicago truncatula PM25 (MtPM25), was proved to be intrinsically disordered and able to prevent aggregation of proteins during freezing, heating, and drying stress treatments [55]. Hydrophobic MtPM25 absorbs up to three-fold more water than MtEM6, a hydrophilic group 1 LEA protein. Although most LEA proteins showed hydrophilic features in their molecular shielding function, the case of MtPM25 showed that different LEA proteins could adopt various stress mechanisms [55]. Besides hydrodynamic volumes, LEA proteins also have an effect on three solvent properties of water (dipolarity/polarizability, hydrogen-bond donor acidity, and hydrogen-bond acceptor basicity), which are involved in one of the proposed in vitro protective mechanisms of these proteins [82]. Post-translational modification such as phosphorylation could regulate the function of LEA proteins by changing their conformations and ligand binding properties [83,84]. For example, phosphorylated salt-tolerance soybean PM18, a member of group 3 LEA proteins, confers better protection of lactate dehydrogenase in the freeze–thaw cycles than its un- or dephosphorylated form [56].
Figure 1 illustrates plant IDPs protecting enzyme activity via a molecular shielding mechanism. The cryoprotection activity seems to not be correlated with specific amino acid sequences or the order of amino acid sequences but rather highly correlated with the biophysical properties of IDPs (i.e., hydrodynamic radius, the density of charged residues) [80,85]. Indeed, the scrambled grape dehydrin peptide showed cryoprotection of lactate dehydrogenase similar to that of the wild-type peptide [80]. Of note, randomly selected IDP fragments from the human genome, whose amino acid sequences are not related to plant dehydrins, also conferred substantial cryoprotection of lactate dehydrogenase [85]. This example emphasized the importance of the disorder nature in the cryoprotection effect. Hence, further examination of enzyme protection activity toward genome-wide plant IDPs will be critical in providing more evidence of their physicochemical properties related to their mechanisms of enzyme cryoprotection.
4. Disorder-to-Order Transition for Membrane Interaction
Membranes are often damaged during freezing and thawing or desiccation and rehydration. The rupture of the plasma membrane is one of the most commonly used indicators of cell death [86]. The group 2 LEA proteins, dehydrins, are often located near the membrane during abiotic stress [87,88,89,90]. They prevent the formation of oxidation-modified lipids and reduce the amount of electrolyte leakage generated by damaged membranes [91]. A recombinant citrus dehydrin prevented peroxidation of soybean liposomes in vitro, and overexpression of this citrus dehydrin enhanced cold tolerance in tobacco [92]. Dehydrins can alleviate oxidative damage in stressed plants by scavenging hydroxyl and peroxyl radicals or binding metals. The major residues in many dehydrins, glycine, histidine, and lysine, may be targets of these radicals [93,94,95]. Malondialdehyde is believed to be the final product of lipid peroxidation in the plant cell membrane. It is an important indicator of membrane system injury and disruption of cellular metabolism [96]. Malondialdehyde and electrolyte leakage in the freezing-tolerant loquat cultivar were found to be related to its higher accumulation of dehydrin proteins during freezing treatment [97]. Overexpression of LEA14 in foxtail millet reduced electrolyte leakage and enhanced salt and drought tolerance [98]. The inhibition of lipid oxidation and electrolyte leakage by LEA proteins and dehydrins help maintain membrane stability and fluidity; the mechanism involved may be found in the membrane binding properties in vitro.
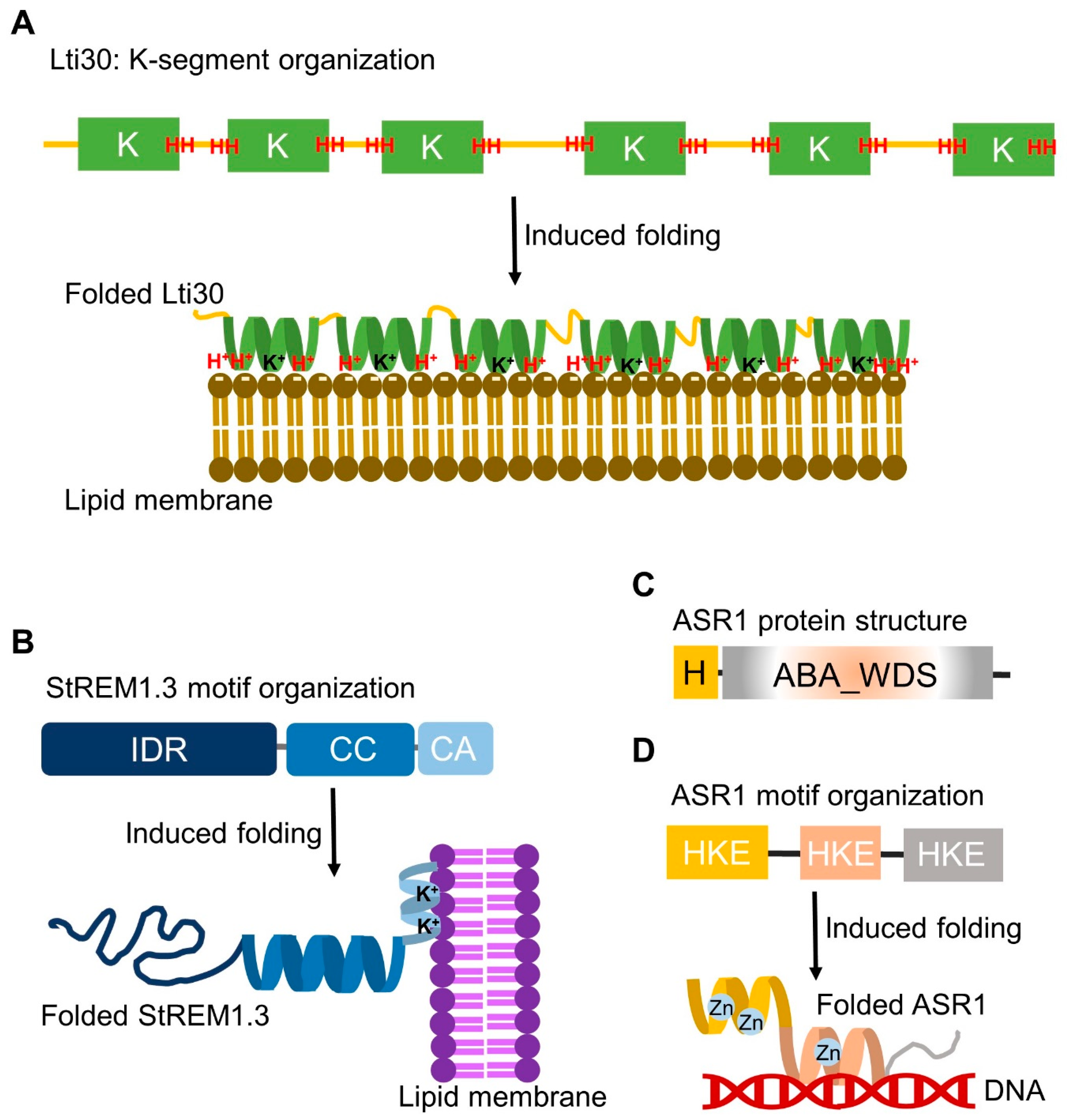
The structural dynamics of dehydrins under stress conditions were summarized [99]. Studies regarding the in vitro interaction of dehydrins with model membranes revealed their disorder-to-order structural transition upon membrane binding [100,101]. This induced folding was shown by disordered K segments of dehydrins gaining ordered α-helix structures upon binding to anionic phospholipid vesicles [102] and micelles [100]. A good example is positively charged Arabidopsis dehydrin Lti30 binding to membranes by recognizing the negatively charged head groups of phospholipids [51] (Figure 2). This membrane interaction decreases the main lipid phase transition by 2.5 °C in vitro [51], corresponding to the decreased survival temperature of 3 °C observed in Arabidopsis overexpressing Lti30 [89]. A nuclear magnetic resonance (NMR) analysis suggested that disordered K segments of Lti30 locally fold into α-helical structures on the membrane surface [101] (Figure 2). Further studies showed that Lti30 may protect the cell by “cross-linking” the membrane lipids [52] and stabilizing the lamellar multilayer structure in response to changing water content [103]. Saltwater cress dehydrins showed induced folding [104], which induced temperature-dependent phase transitions and domain formation of lipid bilayers [105]. Thus, the IDP–membrane interaction may affect conformation changes as well as tertiary and quaternary associations.
IDPs can undergo a disorder-to-order transition and adopt ordered secondary structures such as α-helixes or β-sheets upon environmental changes (high temperature or extreme pH) and in the presence of their binding partners, artificial and natural membranes or osmolytes [6,107]. Besides dehydrins, various LEA proteins undergo the unfolding-to-folding transition to gain ordered conformations in the presence of artificial membranes [83]. Sodium dodecyl sulphate (SDS) is a widely used micelle-forming detergent for generating a membrane-like environment [108]. In the presence of SDS micelles, different groups of LEA proteins adopt α-helical structures; these proteins include AhDHN1 (group 2) from the halophytic species Atriplex halimus [109], pea LEAM (group 3) [110], GmPM1 and GmPM28 (group 4) from soybean [111], GmPM11 (group 1), GmPM6 (group 2), and GmPM30 (group 3) [112]. Other popularly used membrane mimetics are palmitoyl-oleoyl-phosphocholine (POPC) liposomes [113,114]. POPC liposomes induce secondary structure changes of the group 3 LEA protein Arabidopsis LEA7 [115] but not the group 4 LEA protein GmPM1 [54]. The group 3 LEA protein, Arabidopsis COR15A, folds into amphipathic α-helices upon dehydration or in the presence of high-concentration osmolytes [116,117]. COR15A interacts with POPC liposomes at the hydrophobic face of the amphipathic α-helix via hydrophobic interaction [118] and forms oligomeric assemblies that strongly depend on solution osmolarity [119]. The in vitro evidence leads to a hypothesis that COR15A suppresses the formation of non-lamellar lipids, the major cause of membrane damage upon dehydration, and stabilizes cellular membranes to confer freezing tolerance [53,117,118].
The above examples represent the flexible nature of IDPs, whose structural properties are regulated by the binding of a partner (i.e., membrane) as well as the environment (i.e., solution osmolarity). The disorder-to-order conformational change reflects an elegant mechanism of how IDPs might respond rapidly to changing environmental conditions such as stress treatments. Potential implications of the discussed IDP structural dynamics have been proposed as membrane stabilization to protect cells against water deprivation [83]. How conformational changes of IDPs when binding to membranes lead to retention of membrane fluidity and the suppression of membrane fusion events will be the next questions to be answered. Assigning in vivo meaning to in vitro evidence is still challenging.
5. Interactions around the Membrane
The preferred cell membrane localization of dehydrins supports their potential function in membrane protection, such as cold-acclimation-induced dehydrin localization near the cell membranes [88,89,90]. Cell membranes interact with IDPs and also provide an environment for IDPs to interact with their partners. A recent report from BiFC assays showed that pepper dehydrin DHN3 interacts with another dehydrin, HIRD11, most likely at the plasma membrane, but further evidence is required to address the role of this dehydrin–dehydrin interaction in ROS scavenging for drought and salt resistance [120]. The homodimeric and heterodimeric dehydrin–dehydrin interactions can be found in different subcellular distributions. The Arabidopsis dehydrins COR47 and ERD10 formed homodimers in the cytoplasm and RAB18 in the nucleus and cytoplasm [121]. For the moss dehydrin DHNA, the self-association dominates at cellular membranes and is enhanced by high-temperature stress [122]. Moreover, the Arabidopsis dehydrins COR47, ERD10, and RAB18 interact with the aquaporin PIP2B at the plasma membrane [123]. The detailed mechanism of dehydrin–dehydrin interactions and how these interactions affect the interaction of dehydrins with other partners (i.e., aquaporin) await discovery. Autophagy has been reported to regulate the heavy metal stress response [124] and salt [125] and desiccation tolerance via an anti-cell-death function and protein quality control system [20]. Of note, the dehydrin in Medicago truncatula, MtCAS31, interacts with the aquaporin MtPIP2;7 in the ER for autophagic degradation under drought stress [126]. The alfalfa dehydrin DHN1 interacts with two aquaporins, PIP2;1 and TIP1;1, at the plasma membrane [127], but the role of this interaction in controlling intracellular water content against dehydration stress and aluminium toxicity is unknown. Nevertheless, the interaction between dehydrins and aquaporins extends their function from membrane binding to the regulation of membrane proteins. Oligomerization of dehydrins in various cellular compartments may suggest that they have spatial regulation roles for interacting with corresponding membranes of various organelles as well as various partners. Indeed, LEA proteins in Arabidopsis, tomato, and orchid showed wide subcellular distribution [41,60,61].
Remorin is a plant-specific IDP family localized at the plasma membrane and thought to regulate membrane nanodomain assembly [128,129,130]. Potato remorin 1.3 (StREM1.3) is a case of disorder-to-order transition for membrane interaction [130]. The lipid-induced α-helical folding region of the unstructured StREM1.3 C-terminal anchor may interact with the lipid polar heads, whereas the hydrophobic region is embedded inside the lipid phase, forming a nanodomain organization [130]. In comparison to membrane-induced folding of Arabidopsis dehydrin Lti30, both Lti30 and StREM1.3 interact with lipid polar heads through electrostatic interactions via positively charged amino acids (lysine and histidine in Lti30 and lysine in StREM1.3; Figure 2). Although disordered K segments of Lti30 and the C-terminal anchor of StREM1.3 differ in primary amino acid sequences, both undergo lipid-induced folding into α-helical conformations for membrane interactions.
The study of Medicago truncatula remorin SYMREM1 suggested the scaffold role of remorins in regulating plasma membrane nanodomains during the symbiosis process: SYMREM1 stabilizes the interactions between FLOTILLIN 4 and LysM receptor-like kinase LYK3 at the plasma membrane during nodulation-promoting signalling [131]. The phosphorylation of remorins was modulated by biotic and abiotic stressors such as the bacterial “elicitor” peptide Flg22, oligogalacturonides mimicking wounding and herbivory, nitrogen deprivation, ABA, H2O2, cold, osmotic pressure, and salinity [132,133,134,135]. Perception of a virus leads to phosphorylation of group 1 remorins in Solanaceae, thus increasing plasma membrane mobility and facilitating the interaction with remorin-interacting proteins, causing an increase in callose deposition at the plasmodesmata and inhibiting virus cell-to-cell movement [130,136,137,138]. In Arabidopsis, AtREM1.2 and AtREM1.3 interact with general regulatory factor 10 (GRF10) to organize lipid raft nanodomains and participate in plasmodesmata closure to hamper virus spread [66]. The ABA-induced rice remorin OsREM4.1 was associated with OsSERK1 (somatic embryogenesis receptor-like kinase 1) at the plasma membrane, whereas the phosphorylation of OsREM4.1 dissociated the OsREM4.1–OsSERK1 complex; thus, OsSERK1 was able to interact with OsBRI1 (brassinosteroid-insensitive 1) to form the OsBRI1–OsSERK1 receptor kinase complex and activate brassinosteroid (BR) signalling: this process maintains a dynamic equilibrium between ABA and BR signalling [65]. There was a gain of function via the heterologous overexpression of mulberry MiREM1, foxtail millet SiREM6, and poplar PeREM6.5 in Arabidopsis-conferred resistance to salt stress [139,140,141], whereas PeREM6.5 was thought to enhance the plant’s ability to maintain ionic homeostasis under salinity by regulating H+-ATPase activity at the plasma membrane [141].
The disorder nature enables remorins to be an interaction hub scaffolding at the plasma membrane, interacting with various membrane proteins/receptors and thus playing a role in plant–microbe interactions and hormone signalling during stress responses [65,142]. Important topics to be investigated in the future are the functional versatility of remorin family members, consequences of the phosphorylation(s) of remorin-disordered regions in terms of conformational changes and protein–protein interactions, and the molecular mechanisms underlying remorin plasma-membrane–nanodomain organization under different stress conditions. These investigations will provide a deeper understanding of the remorin function in plant interactions with pathogens and symbionts, responses to abiotic stresses, and hormone signalling, which is useful for engineering and modifying remorin-mediated cellular processes for sustainable agriculture.
6. Metal Ion Binding Induced Folding of Stress-Tolerant ASR Proteins
Dehydrins and group 4 LEA proteins showed conformational changes and oligomerization when bound to metal ions, which may function in reducing oxidative damage induced by abiotic stress in plants [143,144,145]. Here, I discuss the group 7 LEA proteins, which correspond to the ASR (ABA, stress, and ripening) protein family that is conserved in many plant species but not found in Arabidopsis [45]. There are 5, 6, and 33 ASR genes in tomato, rice, and wheat, respectively [146,147,148]. Many ASR genes respond to ABA and abiotic stress such as drought, salt, and cold [146,149,150,151,152,153,154,155]. The ASR protein structure consists of a short N-terminal consensus sequence comprising histidine residues and a conserved ABA_WDS (water deficit stress) domain (pfam02496 and IPR003496) in the C-terminal region [156,157]. Heterogeneous expression of ASR genes often confers varying abiotic stress tolerance in different plant species. Overexpressed tomato and wheat ASR1 in tobacco plants increased salt and drought tolerance, respectively [153,158]; overexpression of lily ASR conferred drought and salt tolerance in Arabidopsis by increasing the expression of ABA/stress-regulated genes [159]; and overexpression of ASR from the extreme halophyte Suaeda liaotungensis conferred tolerance to salt, drought, and low temperatures in Arabidopsis [160]. Genetic studies revealed that ASR1-overexpressing tomato plants showed enhanced tolerance to water stress [146,158], and OsASR1-overexpressing rice showed enhanced salinity and drought tolerance [155], but the in vivo mechanism of how ASR proteins enhance stress tolerance is still largely unknown.
Similar to other LEA proteins, ASR proteins are intrinsically disordered. Biochemical and biophysical analyses of ASR proteins provided in vitro data hinting at the potential mechanism involved. Recombinant tomato ASR1 protein possesses zinc-dependent DNA-binding activity [161], which implies its transcription factor function as shown by tomato ASR1 targeting cell-wall-related and aquaporin genes [162]. In vitro studies of recombinant tomato ASR1 showed that ASR1 can adopt different conformations such as α-helix or polyproline type II in response to environmental changes, and the binding of zinc ions promotes α-helix folding and homodimerization of ASR1 [106,163,164]. Disordered segments of tomato ASR1 are enriched with histidine, lysine, and glutamate [106,163]. As compared with the induced folding of dehydrin K segments upon dehydration or membrane binding, the induced folding of tomato ASR1 is due to zinc binding via histidine residues [106,163] (Figure 2). Zinc-induced folding of ASR proteins was also observed in rice, barley, and wheat [165,166]. ASR proteins do not contain any of the known zinc-binding motifs described previously, such as the zinc finger, a common motif in DNA-binding proteins [167]. The abundance of histidine in ASR proteins potentially provides a hint of their possible ability to bind bivalent cations such as zinc [161,168], but direct binding evidence is still lacking. The structural plasticity of ASR proteins in gaining α-helix and β-strand conformations supports their versatile functions; however, the structure–function relationship in stress tolerance awaits investigation. Current data revealed that metal ion binding to ASR proteins induces their folding, but how this conformational change relates to their transactivation capabilities remains unclear. We do not know whether ASR proteins play a role as a chromatin protective protein during stress or as a transcription activator in the stress signal pathway. Future studies using chromatin immunoprecipitation assays will help answer this question.
7. Ectopic IDP Expression Confers Stress Tolerance in Yeast and Bacteria
Stress tolerance phenotypes have been observed in ectopic expression of some plant IDPs in unicellular organisms such as Escherichia coli (E. coli) and yeast. The heterologous hydrophilic domain of LEA expression conferred enhanced desiccation and oxidation tolerance to E. coli [169]. Arabidopsis ERD14 could protect E. coli cells under heat stress [170,171]. Overexpression of cucumber (Cucumis sativus) dehydrin CsLEA11 in E. coli enhanced cell viability and conferred tolerance to heat and cold stress [42]. The expression of eight Arabidopsis LEA genes enhanced desiccation tolerance in yeast [172]. In vivo expression of XsLEAs from the monocot resurrection species Xerophyta schlechteri enhanced E. coli viability under salt, osmotic, and heat stress [57]. Recombinant AcLEA function was evaluated using E. coli as an in vivo model, showing an important protection role against desiccation, oxidant conditions, and osmotic stress [173]. The moss dehydrin DHNA provides stress tolerance to E. coli cells via proteome protection mediated by D-segments [122]. Heterologous expression of ABA_WDS domains from barley and wheat ASR proteins in the yeast Saccharomyces cerevisiae improved its tolerance to salt, heat, and cold stresses [168].
Ectopic expression of plant LEA proteins in unicellular organisms such as E. coli and yeast conferring stress tolerance is probably due to the widely distributed properties of hydrophilins across kingdoms. However, plant IDPs other than hydrophilins also confer stress tolerance in unicellular organisms. For example, a plant-specific DNA binding with one finger (DOF) transcription factor, rice DOF27, is intrinsically disordered and is implicated in the thermotolerance response in yeast [174]. A member from the Poaceae-specific RICE SALT SENSITIVE 1 (RSS1) family, unstructured wheat RSS1-Like 1, enhanced stress tolerance of Arabidopsis seedlings and yeast cells [175,176]. Therefore, ectopic expression of plant IDPs in protecting cellular matter across multiple kingdoms of life may not simply be attributed to the effect of the homology. The principle of physiochemical properties governing plant IDPs underpinning their quick sensitivity and responsiveness to changing environments and dynamic conformations to interact with various partners for stress tolerance needs to be investigated. Given that tardigrade disordered proteins are proposed to undergo LLPS and interact with various partners for transitioning into an anhydrobiotic status under extreme conditions [177], LLPS is potentially a way for plant IDPs to change the physiological status in different organisms to achieve an anhydrobiosis-like status for stress resistance.
8. Roles of LLPS in Plant Stress Responses
Disordered proteins have phase transition properties for undergoing LLPS and forming membrane-less biomolecular condensates. These properties enable a rapid assembly, disassembly, and concentration of cellular components and facilitate the dynamic formation of local reaction centres with spatiotemporal specificity [27,178]. Notably, LLPS is also involved in plant developmental phase transition such as flowering and seed germination [11]. The FLOWERING CONTROL LOCUS A (FCA) nuclear body driven by highly disordered FLX-LIKE 2 (FLL2) plays a role in transcriptional regulation to control flowering [179]. Stress granules are among the well-studied membrane-less condensates transiently assembled via LLPS in response to stress [180,181], whereas LLPS is induced when plants encounter various abiotic stresses such as a high temperature (35 °C and 37 °C) [182,183], a low temperature (4 °C) [184], salinity (0.4 M NaCl treatment) [185], and ROS (1 mM H2O2) [186]. LLPS driven by the IDR of phytochrome B (phyB) plays a major role in temperature sensing and thermomorphogenesis, suggesting an emerging mechanism for plants to directly respond to thermal changes [187]. A plant-specific prion-like protein was named FLOE1, the definition of floe being “a sheet of floating ice”, which is a phase-separated body of water [188]. Upon hydration, LLPS of intrinsically disordered FLOE1 allowed the embryo to sense water stress and promoted seed germination [188]. The importance of LLPS in the osmotic stress response was shown by the Arabidopsis transcriptional regulator SEUSS (SEU). The IDR of SEU is responsible for forming liquid-like nuclear condensates, which is indispensable for osmotic stress tolerance [189]. Unlike the well-established mechanism of FCA nuclear condensates in transcriptional control of flowering, we do not know whether the components of SEU nuclear condensates contain multiple active transcription regulators for promoting the transcription of osmotic stress-responsive genes.
Besides the aforementioned abiotic stress responses, LLPS is also involved in the biotic stress response as an interacting hub in plant immunity. LLPS is induced by bacterial phytopathogens Pseudomonas syringae pv. Maculicola ES4326 and P. syringae pv. tomato DC3000 as shown by the case of guanylate-binding protein (GBP)-like GTPases (GBPLs) [67]. The IDR of GBPLs is required for the assembly of LLPS-driven condensates within the nucleus and they are called GBPL defence-activated condensates, which coordinate defence-gene transcription for disease resistance [67]. The IDR of the salicylic acid receptor NONEXPRESSOR-OF-PATHOGENESIS-RELATED GENE 1 (NPR1) forms cytoplasmic condensates and serves as an interacting hub to sequester and degrade proteins involved in programmed cell death in infection-adjacent cells with a low pathogen load [30]. LLPS of NPR1 plays a role in switching cellular programming from effector-triggered immunity (ETI) in infected cells to systemic acquired resistance in adjacent cells [30]. A recent report highlighted the importance of LLPS in regulating translational reprogramming in plant immunity [32]. Arabidopsis HEM1 (named for heme biosynthesis in the yeast homologue) contains a plant-specific IDR called the low-complexity domain, which undergoes LLPS to form condensates to interact with translation factors, thus suppressing translation efficiency of the pro-death immune genes during ETI [32]. Although the interaction of HEM1 with NPR1 in the cytoplasmic condensates has not been detected [32], both cases suggest a common role of LLPS in balancing tissue health and disease resistance during the plant immune response.
Protein disorder has a versatile contribution to both the plant immune system and pathogen virulence [190,191]. The N-terminal IDR of prosystemin, the 200-aa precursor of the peptidic hormone systemin responding to wounding and herbivore attack, was able to induce defence-related genes and protect tomato plants against Botrytis cinerea and Spodoptera littoralis larvae [192]. In the plant defence response, RPM1-INTERACTING PROTEIN 4 (RIN4) is a negative regulator and targeted by multiple bacteria effectors [193,194,195]. The IDR of RIN4 was known to fold various conformations when interacting with different partners such as bacterial effectors and be under regulation of post-translational modification [196,197,198,199]. The latest report suggested that the intrinsically disordered nature of RIN4 provides a flexible platform to broaden pathogen recognition specificity by establishing compatibility with otherwise incompatible leucine-rich repeat immune receptor proteins [200,201]. Besides its involvement in plant immunity responses, LLPS is involved in pathogen virulence. The IDR of FgRpd3 (reduced potassium dependency 3) from wheat head blight fungus Fusarium graminearumis undergoes LLPS, which assists its interaction with inhibitor of growth (ING) proteins for regulating histone deacetylation and gene expression, thus affecting fungal development and pathogenicity [202].
Achieving the necessary spatiotemporal resolution to deduce the parameters that govern the assembly and behaviour of LLPS is challenging [203], especially when it involves the dynamics and multiple subcellular locations of IDPs (i.e., LEA proteins). Therefore, as compared with extensive studies of plant LEA proteins in stress tolerance, much less research has investigated plant LEA proteins promoting LLPS [204]. Biomolecular condensates formed by LEA proteins from the anhydrobiotic animal brine shrimp were considered to act as protective compartments for desiccation-sensitive proteins to promote dehydration tolerance during anhydrobiosis [205,206]. Whether LLPS has a similar role in plant LEA protein functioning in stress tolerance still awaits investigation. Despite recent discoveries of novel plant IDPs involved in LLPS to generate biomolecular condensates during stress sensing and signalling, several questions remain. What are the determining factors of LLPS for forming biomolecular condensates (i.e., specific amino acid sequences of IDPs, specific IDP conformation, or biophysical properties)? And what are the components of biomolecular condensates in various stress situations? Future investigation dissecting the spatial and temporal rules guiding biomolecular condensate formation under various stress conditions is important to understand the roles of plant IDP-driven LLPS in stress responses.
9. IDPs Interacting with the Cytoskeleton for Stress Adaptation
Changes at the cytoskeleton protein level and dynamics were observed upon cold [207], drought [208], and heavy metal [209] stress [210]. Depolymerization of actin filaments may prevent expansion-induced cell lysis during freeze−thaw cycles [211], and the cytoskeleton was thought to take centre stage in abiotic stress responses and resilience [212]. The rapid remodelling of the cytoskeleton can coordinate the movement of intracellular organelles, formation of immune microdomain complexes, transportation of defence compounds and turnover of recognizing receptors, cell-wall-based defence upon pathogen attack, as well as hormone-mediated cell expansion and division under abiotic stress [213,214,215,216,217]. Many cytoskeleton-associated proteins are IDPs or possess extensive IDRs [218,219]. The disordered nature of these cytoskeleton-associated proteins enables them to regulate the dynamics and organization of highly ordered cytoskeleton proteins for quick responsiveness to the changing environment. Dehydrins are thought to be able to stabilize the cytoskeleton under stress conditions, as shown by dehydrins from Thellungiella salsuginea facilitating the polymerization of actin filaments [211]. Recent findings highlighted the importance of IDPs in interacting with the cytoskeleton for stress adaptation in plants. Rice intrinsically disordered repetitive pro-rich protein (RePRP) interacts with both the actin and tubulin cytoskeleton to generate short-but-heavy roots under a water deficit [64]. Arabidopsis COMPANION OF CELLULOSE SYNTHASE1 (CC1) controls microtubule bundling and dynamics to sustain plant growth under salt stress via its intrinsically disordered N-terminus, which can diffuse bidirectionally along the microtubule lattice [220], similar to the mammalian tubulin-associated unit Tau [221]. As compared with the well-established mechanism of disordered Tau-microtubule interaction causing neurodegeneration, how protein disorders regulate cytoskeletal dynamics in plant stress responses is unclear. We still lack studies investigating why protein disorder interacting with the cytoskeleton results in neuron degenerative disease in mammals but stress adaptation in plants. Treatment with the actin depolymerizing drug latrunculin B can destabilize the association of SYMREM1 to the plasma membrane nanodomains [131], so the cytoskeleton may be involved in remorin scaffolding. In fact, both rice and Arabidopsis remorins interact with the actin cytoskeleton in plants [222,223,224]. The IDR of Arabidopsis remorin1.2 orchestrates high-order self-assembly and formin clustering to mediate a nanocluster of membrane nanodomains for actin remodelling during innate immune responses [224]. Such molecular condensation-regulated cytoskeleton remodelling has been identified in the mammalian system [225]. The similarity and differences of protein disorder mediating cytoskeleton dynamics between plant and animal systems need to be further investigated. Future research investigating the mechanism of molecular condensation regulating the cell wall–plasma membrane–cytoskeleton continuum during plant immune responses will answer the question of how protein disorder affects stress sensing and signalling. Because of the continuing race of disorder between plant hosts and pathogens, not surprisingly, a bacteria effector hijacks the host actin cytoskeleton, as shown by the Xanthomonas effector XopR affecting actin nucleation, crosslinking actin filaments and actin depolymerization in host cells via IDR-mediated LLPS [226]. Thus, IDP/IDR-mediated cytoskeleton dynamics are a double-edged sword in the biotic stress response in that they can affect membrane nanodomain distribution for plant defence (e.g., the case of remorins) and can be a hijacked target of pathogen infection.
10. Conclusions and Perspectives
The plasticity of IDPs is an easy and fast way for sessile plants to introduce versatility into protein interaction networks and quickly and efficiently adapt to environmental changes [227]. The ability of IDPs to interact with various partners gives them versatile functions in different cellular processes at various subcellular compartments during stress (Figure 3). With their multiple roles explored, the next step is to apply modern technologies in cell biology and biophysics for understanding the underlying mechanism [228,229,230,231,232]. Bioimaging tools are useful for studying plant physiological processes in vivo [233]. A recent report suggested that fixation causes artifacts of LLPS in the animal system [234], so live-cell imaging could be a preferable approach to dissect the phenomenon of LLPS. Single-particle tracking (SPT) is a valuable method for directly measuring and calculating the diffusion and dynamics within biomolecular condensates [203]. A recent study using SPT in yeast cells revealed the mechanism of mobility of RAD52 (named radiation-sensitive) within repair foci at DNA double-stranded break events and suggested that LLPS was required for DNA repair [235]. In plants, an SPT method has been developed for studying the distribution and dynamics of the plant membrane protein aquaporin [236]. Although not reported so far, using SPT with super-resolution microscopy is expected to dissect the movement of plant IDPs and the spatial and temporal regulation of LLPS (i.e., the formation of biomolecular condensates) during stress responses by directly “observing” how they diffuse in living cells in front of our eyes.
Of note, the fluorescence tags used in live-cell technology may alter LLPS and IDP properties, potentially causing artifacts [237,238]. Therefore, biophysics approaches can be complementary to bioimaging tools for overcoming these obstacles. Biophysics characterization such as NMR spectroscopy can describe the binding mechanisms and map the conformational dynamics of IDPs [228,239], and in-cell NMR is a promising solution to provide their structural information in living cells [240]. A preliminary study using in-cell NMR in bacteria showed the disordered nature of Arabidopsis dehydrin ERD14 in the cytoplasm of E. coli [170]. Future research to investigate the conformational changes in ERD14 upon stress treatments via in-cell NMR will help answer the question of how ectopic expression of plant IDPs confers stress tolerance in bacteria. In plants, the current bottleneck of in-cell NMR is the delivery of isotope-labelled proteins into living plant cells, but exogenous agents (bacteria and viruses) and cell-penetrating peptides could be used for eliminating the bottleneck [241]. Taken together, future research should use multidisciplinary approaches, especially combined with cell biology and biophysics tools, to catch the dynamic motions of plant IDPs both in vitro and in vivo.
Plant stress-responsive IDPs can have broad application beyond plant research, based on basic research of IDPs in plant stress responses as mentioned previously. A recent innovation showed that a FRET biosensor can monitor rapid intracellular changes under osmotic stress by utilizing the environmental sensitivity and structural plasticity of Arabidopsis LEA4-5 protein [242]. A better understanding of the biochemical properties and conformational information of plant IDPs under various stress conditions will provide the knowledge needed for programming, such as the amino acid sequence and responsiveness profiling. We should be able to produce synthetic peptides based on plant IDPs for different applications. For example, LEA proteins have promising cryoprotection properties, preserve desiccated biomaterials, and can act as anti-osmotic agents, which leads to novel opportunities in biomedical and pharmacological development [243].
With an increasing human population, food security is certainly one of the most serious global challenges today. Because IDPs have multiple functions in protecting plants under various abiotic and biotic stress conditions, plant stress-responsive IDPs have potential application as genetic engineering targets for generating stress-resistant crop plants. A potential future research direction is specifically targeting stress-responsive LLPS processes in crop plants to achieve temporary stress resistance during environmentally harsh periods. Understanding the mechanisms used by these IDPs that allow them to mediate abiotic and biotic tolerance in plants will not only close the knowledge gap but also reveal insights that may revolutionize crop resilience. Using multidisciplinary technologies to unravel the intricate workings of IDPs in plant stress adaptation will pave the way for the development of more robust and sustainable crops to solve food security problems.
Funding
A.-S.H. was supported by a Royal Society grant and a Gatsby Charitable Foundation grant led by Michael Webster.
Acknowledgments
Icons in Figure 3 were designed using macrovector_official/Freepik (http://www.freepik.com) (accessed on 28 October 2023). The author thanks all the reviewers and editors for their constructive suggestions to improve this manuscript.
Conflicts of Interest
The author declares no conflict of interest.
References
- Dilmore, R.; Zhang, L. Greenhouse gases and their role in climate change. In Green Energy and Technology; Springer: Berlin/Heidelberg, Germany, 2018; pp. 15–32. [Google Scholar]
- Zandalinas, S.I.; Fritschi, F.B.; Mittler, R. Global warming, climate change, and environmental pollution: Recipe for a multifactorial stress combination disaster. Trends Plant Sci. 2021, 26, 588–599. [Google Scholar] [CrossRef] [PubMed]
- Bailey-Serres, J.; Parker, J.E.; Ainsworth, E.A.; Oldroyd, G.E.D.; Schroeder, J.I. Genetic strategies for improving crop yields. Nature 2019, 575, 109–118. [Google Scholar] [PubMed]
- Zhang, H.; Zhu, J.; Gong, Z.; Zhu, J.K. Abiotic stress responses in plants. Nat. Rev. Genet. 2022, 23, 104–119. [Google Scholar]
- Waadt, R.; Seller, C.A.; Hsu, P.K.; Takahashi, Y.; Munemasa, S.; Schroeder, J.I. Plant hormone regulation of abiotic stress responses. Nat. Rev. Mol. Cell Biol. 2022, 23, 680–694. [Google Scholar] [CrossRef] [PubMed]
- Uversky, V.N. Intrinsically disordered proteins and their environment: Effects of strong denaturants, temperature, pH, counter ions, membranes, binding partners, osmolytes, and macromolecular crowding. Protein J. 2009, 28, 305–325. [Google Scholar] [PubMed]
- Uversky, V.N. Intrinsically disordered proteins and their “mysterious” (meta)physics. Front. Phys. 2019, 7, 10. [Google Scholar] [CrossRef]
- Tompa, P.; Schad, E.; Tantos, A.; Kalmar, L. Intrinsically disordered proteins: Emerging interaction specialists. Curr. Opin. Struct. Biol. 2015, 35, 49–59. [Google Scholar]
- Wright, P.E.; Dyson, H.J. Intrinsically disordered proteins in cellular signalling and regulation. Nat. Rev. Mol. Cell Biol. 2015, 16, 18–29. [Google Scholar]
- Dirk, L.M.A.; Abdel, C.G.; Ahmad, I.; Neta, I.C.S.; Pereira, C.C.; Pereira, F.E.C.B.; Unêda-Trevisoli, S.H.; Pinheiro, D.G.; Downie, A.B. Late embryogenesis abundant protein-client protein interactions. Plants 2020, 9, 814. [Google Scholar]
- Hsiao, A.S. Plant protein disorder: Spatial regulation, broad specificity, switch of signaling and physiological status. Front. Plant Sci. 2022, 13, 904446. [Google Scholar]
- Covarrubias, A.A.; Romero-Pérez, P.S.; Cuevas-Velazquez, C.L.; Rendón-Luna, D.F. The functional diversity of structural disorder in plant proteins. Arch. Biochem. Biophys. 2020, 680, 108229. [Google Scholar]
- Bondos, S.E.; Dunker, A.K.; Uversky, V.N. Intrinsically disordered proteins play diverse roles in cell signaling. Cell Commun. Signal. 2022, 20, 20. [Google Scholar] [PubMed]
- Cuevas-Velazquez, C.L.; Rendón-Luna, D.F.; Covarrubias, A.A. Dissecting the cryoprotection mechanisms for dehydrins. Front. Plant Sci. 2014, 5, 583. [Google Scholar] [CrossRef] [PubMed]
- Kosová, K.; Vítámvás, P.; Prášil, I.T. Wheat and barley dehydrins under cold, drought, and salinity—What can LEA-II proteins tell us about plant stress response? Front. Plant Sci. 2014, 5, 343. [Google Scholar] [CrossRef] [PubMed]
- Covarrubias, A.A.; Cuevas-Velazquez, C.L.; Romero-Pérez, P.S.; Rendón-Luna, D.F.; Chater, C.C.C. Structural disorder in plant proteins: Where plasticity meets sessility. Cell. Mol. Life Sci. 2017, 74, 3119–3147. [Google Scholar] [PubMed]
- Boothby, T.C.; Tapia, H.; Brozena, A.H.; Piszkiewicz, S.; Smith, A.E.; Giovannini, I.; Rebecchi, L.; Pielak, G.J.; Koshland, D.; Goldstein, B. Tardigrades use intrinsically disordered proteins to survive desiccation. Mol. Cell 2017, 65, 975–984.e5. [Google Scholar] [CrossRef] [PubMed]
- Arakawa, K.; Numata, K. Reconsidering the “glass transition” hypothesis of intrinsically unstructured CAHS proteins in desiccation tolerance of tardigrades. Mol. Cell. 2021, 81, 409–410. [Google Scholar] [PubMed]
- Zamora-Briseño, J.A.; Pereira-Santana, A.; Reyes-Hernández, S.J.; Cerqueda-García, D.; Castaño, E.; Rodríguez-Zapata, L.C. Towards an understanding of the role of intrinsic protein disorder on plant adaptation to environmental challenges. Cell Stress Chaperones 2021, 26, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Mladenov, P.; Zasheva, D.; Planchon, S.; Leclercq, C.C.; Falconet, D.; Moyet, L.; Brugière, S.; Moyankova, D.; Tchorbadjieva, M.; Ferro, M.; et al. Proteomics evidence of a systemic response to desiccation in the resurrection plant Haberlea rhodopensis. Int. J. Mol. Sci. 2022, 23, 8520. [Google Scholar] [CrossRef]
- Choura, M.; Rebaï, A.; Hanin, M. Proteome-wide analysis of protein disorder in Triticum aestivum and Hordeum vulgare. Comput. Biol. Chem. 2020, 84, 107138. [Google Scholar] [CrossRef]
- Sun, X.; Rikkerink, E.H.; Jones, W.T.; Uversky, V.N. Multifarious roles of intrinsic disorder in proteins illustrate its broad impact on plant biology. Plant Cell 2013, 25, 38–55. [Google Scholar]
- Már, M.; Nitsenko, K.; Heidarsson, P.O. Multifunctional Intrinsically Disordered Regions in Transcription Factors. Chemistry 2023, 29, e202203369. [Google Scholar] [CrossRef]
- Reinar, W.B.; Greulich, A.; Stø, I.M.; Knutsen, J.B.; Reitan, T.; Tørresen, O.K.; Jentoft, S.; Butenko, M.A.; Jakobsen, K.S. Adaptive protein evolution through length variation of short tandem repeats in Arabidopsis. Sci. Adv. 2023, 9, eadd6960. [Google Scholar] [CrossRef]
- Haynes, C.; Oldfield, C.J.; Ji, F.; Klitgord, N.; Cusick, M.E.; Radivojac, P.; Uversky, V.N.; Vidal, M.; Iakoucheva, L.M. Intrinsic disorder is a common feature of hub proteins from four eukaryotic interactomes. PLoS Comput. Biol. 2006, 2, e100. [Google Scholar]
- Cuevas-Velazquez, C.L.; Dinneny, J.R. Organization out of disorder: Liquid-liquid phase separation in plants. Curr. Opin. Plant Biol. 2018, 45, 68–74. [Google Scholar] [PubMed]
- Kim, J.; Lee, H.; Lee, H.G.; Seo, P.J. Get closer and make hotspots: Liquid-liquid phase separation in plants. EMBO Rep. 2021, 22, e51656. [Google Scholar] [CrossRef]
- Hirose, T.; Ninomiya, K.; Nakagawa, S.; Yamazaki, T. A guide to membraneless organelles and their various roles in gene regulation. Nat. Rev. Mol. Cell Biol. 2023, 24, 288–304. [Google Scholar] [CrossRef] [PubMed]
- Boccaccio, G.L.; Thomas, M.G.; García, C.C. Membraneless organelles and condensates orchestrate innate immunity against viruses. J. Mol. Biol. 2023, 435, 167976. [Google Scholar] [CrossRef]
- Zavaliev, R.; Mohan, R.; Chen, T.; Dong, X. Formation of NPR1 condensates promotes cell survival during the plant immune response. Cell 2020, 182, 1093–1108. [Google Scholar] [PubMed]
- Wang, H.; Niu, R.; Zhou, Y.; Tang, Z.; Xu, G.; Zhou, G. ECT9 condensates with ECT1 and regulates plant immunity. Front. Plant Sci. 2023, 14, 1140840. [Google Scholar]
- Zhou, Y.; Niu, R.; Tang, Z.; Mou, R.; Wang, Z.; Zhu, S.; Yang, H.; Ding, P.; Xu, G. Plant HEM1 specifies a condensation domain to control immune gene translation. Nat. Plants 2023, 9, 289–301. [Google Scholar] [CrossRef]
- Field, S.; Jang, G.J.; Dean, C.; Strader, L.C.; Rhee, S.Y. Plants use molecular mechanisms mediated by biomolecular condensates to integrate environmental cues with development. Plant Cell 2023, 35, 3173–3186. [Google Scholar]
- Boothby, T.C.; Pielak, G.J. Intrinsically disordered proteins and desiccation tolerance: Elucidating functional and mechanistic underpinnings of anhydrobiosis. Bioessays 2017, 39, 1700119. [Google Scholar]
- Tunnacliffe, A.; Wise, M.J. The continuing conundrum of the LEA proteins. Naturwissenschaften 2007, 94, 791–812. [Google Scholar]
- Leprince, O.; Pellizzaro, A.; Berriri, S.; Buitink, J. Late seed maturation: Drying without dying. J. Exp. Bot. 2017, 68, 827–841. [Google Scholar] [CrossRef]
- Hincha, D.K.; Thalhammer, A. LEA proteins: IDPs with versatile functions in cellular dehydration tolerance. Biochem. Soc. Trans. 2012, 40, 1000–1003. [Google Scholar] [CrossRef]
- Wang, X.S.; Zhu, H.B.; Jin, G.L.; Liu, H.L.; Wu, W.R.; Zhu, J. Genome-scale identification and analysis of LEA genes in rice (Oryza sativa L.). Plant Sci. 2007, 172, 414–420. [Google Scholar] [CrossRef]
- Jia, C.; Guo, B.; Wang, B.; Li, X.; Yang, T.; Li, N.; Wang, J.; Yu, Q. The LEA gene family in tomato and its wild relatives: Genome-wide identification, structural characterization, expression profiling, and role of SlLEA6 in drought stress. BMC Plant Biol. 2022, 22, 596. [Google Scholar] [CrossRef] [PubMed]
- Hundertmark, M.; Hincha, D.K. LEA (late embryogenesis abundant) proteins and their encoding genes in Arabidopsis thaliana. BMC Genom. 2008, 9, 118. [Google Scholar] [CrossRef]
- Cao, J.; Li, X. Identification and phylogenetic analysis of late embryogenesis abundant proteins family in tomato (Solanum lycopersicum). Planta 2015, 241, 757–772. [Google Scholar] [CrossRef]
- Artur, M.A.S.; Zhao, T.; Ligterink, W.; Schranz, E.; Hilhorst, H.W.M. Dissecting the genomic diversification of late embryogenesis abundant (LEA) protein gene families in plants. Genome Biol. Evol. 2019, 11, 459–471. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Li, G.-J.; Bressan, R.A.; Song, C.-P.; Zhu, J.-K.; Zhao, Y. Abscisic acid dynamics, signaling, and functions in plants. J. Integr. Plant Biol. 2020, 62, 25–54. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Li, S.; Chen, W.; Zhang, J.; Zhang, L.; Sun, W.; Wang, Z. Plant dehydrins: Expression, regulatory networks, and protective roles in plants challenged by abiotic stress. Int. J. Mol. Sci. 2021, 22, 12619. [Google Scholar] [CrossRef] [PubMed]
- Battaglia, M.; Olvera-Carrillo, Y.; Garciarrubio, A.; Campos, F.; Covarrubias, A.A. The enigmatic LEA proteins and other hydrophilins. Plant Physiol. 2008, 148, 6–24. [Google Scholar] [CrossRef] [PubMed]
- Chakrabortee, S.; Boschetti, C.; Walton, L.J.; Sarkar, S.; Rubinsztein, D.C.; Tunnacliffe, A. Hydrophilic protein associated with desiccation tolerance exhibits broad protein stabilization function. Proc. Natl. Acad. Sci. USA 2007, 104, 18073–18078. [Google Scholar] [CrossRef]
- Kim, S.X.; Çamdere, G.; Hu, X.; Koshland, D.; Tapia, H. Synergy between the small intrinsically disordered protein Hsp12 and trehalose sustain viability after severe desiccation. Elife 2018, 7, e38337. [Google Scholar]
- Koshland, D.; Tapia, H. Desiccation tolerance: An unusual window into stress biology. Mol. Biol. Cell 2019, 30, 737–741. [Google Scholar] [CrossRef]
- Abdul Aziz, M.; Sabeem, M.; Mullath, S.K.; Brini, F.; Masmoudi, K. Plant group II LEA proteins: Intrinsically disordered structure for multiple functions in response to environmental stresses. Biomolecules 2021, 11, 1662. [Google Scholar]
- Kovacs, D.; Kalmar, E.; Torok, Z.; Tompa, P. Chaperone activity of ERD10 and ERD14, two disordered stress-related plant proteins. Plant Physiol. 2008, 147, 381–390. [Google Scholar] [CrossRef]
- Eriksson, S.K.; Kutzer, M.; Procek, J.; Gröbner, G.; Harryson, P. Tunable membrane binding of the intrinsically disordered dehydrin Lti30, a cold-induced plant stress protein. Plant Cell 2011, 23, 2391–2404. [Google Scholar] [CrossRef]
- Gupta, A.; Marzinek, J.K.; Jefferies, D.; Bond, P.J.; Harryson, P.; Wohland, T. The disordered plant dehydrin Lti30 protects the membrane during water-related stress by cross-linking lipids. J. Biol. Chem. 2019, 294, 6468–6482. [Google Scholar] [CrossRef] [PubMed]
- Steponkus, P.L.; Uemura, M.; Joseph, R.A.; Gilmour, S.J.; Thomashow, M.F. Mode of action of the COR15a gene on the freezing tolerance of Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 1998, 95, 14570–14575. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Sun, Y.; Liu, Y.; Zou, Y.; Huang, J.; Zheng, Y.; Liu, G. The N-terminal region of soybean PM1 protein protects liposomes during freeze-thaw. Int. J. Mol. Sci. 2020, 21, 5552. [Google Scholar] [PubMed]
- Boucher, V.; Buitink, J.; Lin, X.; Boudet, J.; Hoekstra, F.A.; Hundertmark, M.; Renard, D.; Leprince, O. MtPM25 is an atypical hydrophobic late embryogenesis-abundant protein that dissociates cold and desiccation-aggregated proteins. Plant Cell Environ. 2010, 33, 418–430. [Google Scholar] [CrossRef]
- Liu, Y.; Yang, M.; Cheng, H.; Sun, N.; Liu, S.; Li, S.; Wang, Y.; Zheng, Y.; Uversky, V.N. The effect of phosphorylation on the salt-tolerance-related functions of the soybean protein PM18, a member of the group-3 LEA protein family. Biochim. Biophys. Acta Proteins Proteom. 2017, 1865, 1291–1303. [Google Scholar]
- Artur, M.A.S.; Rienstra, J.; Dennis, T.J.; Farrant, J.M.; Ligterink, W.; Hilhorst, H. Structural plasticity of intrinsically disordered LEA proteins from Xerophyta schlechteri provides protection in vitro and in vivo. Front. Plant Sci. 2019, 10, 1272. [Google Scholar] [CrossRef] [PubMed]
- Lv, A.; Su, L.; Wen, W.; Fan, N.; Zhou, P.; An, Y. Analysis of the function of the alfalfa Mslea-D34 gene in abiotic stress responses and flowering time. Plant Cell Physiol. 2021, 62, 28–42. [Google Scholar] [CrossRef]
- Lv, A.; Su, L.; Liu, X.; Xing, Q.; Huang, B.; An, Y.; Zhou, P. Characterization of dehydrin protein, CdDHN4-L and CdDHN4-S, and their differential protective roles against abiotic stress in vitro. BMC Plant Biol. 2018, 18, 299. [Google Scholar] [CrossRef]
- Candat, A.; Paszkiewicz, G.; Neveu, M.; Gautier, R.; Logan, D.C.; Avelange-Macherel, M.H.; Macherel, D. The ubiquitous distribution of late embryogenesis abundant proteins across cell compartments in Arabidopsis offers tailored protection against abiotic stress. Plant Cell 2014, 26, 3148–3166. [Google Scholar] [CrossRef]
- Ling, H.; Zeng, X.; Guo, S. Functional insights into the late embryogenesis abundant (LEA) protein family from Dendrobium officinale (Orchidaceae) using an Escherichia coli system. Sci. Rep. 2016, 6, 39693. [Google Scholar] [CrossRef]
- Liu, J.; Liu, J.; Deng, L.; Liu, H.; Liu, H.; Zhao, W.; Zhao, Y.; Sun, X.; Fan, S.; Wang, H.; et al. An intrinsically disordered region-containing protein mitigates the drought-growth trade-off to boost yields. Plant Physiol. 2023, 192, 274–292. [Google Scholar] [CrossRef]
- Perochon, A.; Jianguang, J.; Kahla, A.; Arunachalam, C.; Scofield, S.R.; Bowden, S. TaFROG encodes a Pooideae orphan protein that Interacts with SnRK1 and enhances resistance to the mycotoxigenic fungus Fusarium graminearum. Plant Physiol. 2015, 169, 2895–2906. [Google Scholar] [CrossRef]
- Hsiao, A.S.; Wang, K.; Ho, T.D. An intrinsically disordered protein interacts with the cytoskeleton for adaptive root growth under stress. Plant Physiol. 2020, 183, 570–587. [Google Scholar] [CrossRef]
- Gui, J.; Zheng, S.; Liu, C.; Shen, J.; Li, J.; Li, L. OsREM4.1 interacts with OsSERK1 to coordinate the interlinking between abscisic acid and brassinosteroid signaling in rice. Dev. Cell 2016, 38, 201–213. [Google Scholar] [CrossRef]
- Huang, D.; Sun, Y.; Ma, Z.; Ke, M.; Cui, Y.; Chen, Z.; Chen, C.; Ji, C.; Tran, T.M.; Yang, L.; et al. Salicylic acid-mediated plasmodesmal closure via Remorin-dependent lipid organization. Proc. Natl. Acad. Sci. USA 2019, 116, 21274–21284. [Google Scholar] [PubMed]
- Huang, S.; Zhu, S.; Kumar, P.; MacMicking, J.D. A phase-separated nuclear GBPL circuit controls immunity in plants. Nature 2021, 594, 424–429. [Google Scholar] [PubMed]
- Petersen, J.; Eriksson, S.K.; Harryson, P.; Pierog, S.; Colby, T.; Bartels, D.; Röhrig, H. The lysine-rich motif of intrinsically disordered stress protein CDeT11-24 from Craterostigma plantagineum is responsible for phosphatidic acid binding and protection of enzymes from damaging effects caused by desiccation. J. Exp. Bot. 2012, 63, 4919–4929. [Google Scholar]
- Popova, A.V.; Rausch, S.; Hundertmark, M.; Gibon, Y.; Hincha, D.K. The intrinsically disordered protein LEA7 from Arabidopsis thaliana protects the isolated enzyme lactate dehydrogenase and enzymes in a soluble leaf proteome during freezing and drying. Biochim. Biophys. Acta 2015, 1854, 1517–1525. [Google Scholar] [PubMed]
- Zhou, Y.; He, P.; Xu, Y.; Liu, Q.; Yang, Y.; Liu, S. Overexpression of CsLEA11, a Y3SK2-type dehydrin gene from cucumber (Cucumis sativus), enhances tolerance to heat and cold in Escherichia coli. AMB Express 2017, 7, 182. [Google Scholar] [CrossRef] [PubMed]
- Rendón-Luna, D.F.; Romero-Pérez, P.S.; Cuevas-Velazquez, C.L.; Reyes, J.L.; Covarrubias, A.A. Determining the protective activity of IDPs under partial dehydration and freeze-yhaw conditions. Methods Mol. Biol. 2020, 2141, 519–528. [Google Scholar] [PubMed]
- Yokoyama, T.; Ohkubo, T.; Kamiya, K.; Hara, M. Cryoprotective activity of Arabidopsis KS-type dehydrin depends on the hydrophobic amino acids of two active segments. Arch. Biochem. Biophys. 2020, 691, 108510. [Google Scholar] [CrossRef]
- Singh, K.K.; Graether, S.P. The in vitro structure and functions of the disordered late embryogenesis abundant three proteins. Protein Sci. 2021, 30, 678–692. [Google Scholar] [CrossRef] [PubMed]
- Tan, F.; Sun, N.; Zhang, L.; Wu, J.; Xiao, S.; Tan, Q.; Uversky, V.N.; Liu, Y. Functional characterization of an unknown soybean intrinsically disordered protein in vitro and in Escherichia coli. Int. J. Biol. Macromol. 2021, 166, 538–549. [Google Scholar] [CrossRef]
- Abdul Aziz, M.; Sabeem, M.; Kutty, M.S.; Rahman, S.; Alneyadi, M.K.; Alkaabi, A.B.; Almeqbali, E.S.; Brini, F.; Vijayan, R.; Masmoudi, K. Enzyme stabilization and thermotolerance function of the intrinsically disordered LEA2 proteins from date palm. Sci. Rep. 2023, 13, 11878. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Qin, J.; Chen, M.; Sun, N.; Tan, F.; Zhang, H.; Zou, Y.; Uversky, V.N.; Liu, Y. The moonlighting function of soybean disordered methyl-CpG-binding domain 10c protein. Int. J. Mol. Sci. 2023, 24, 8677. [Google Scholar]
- Goyal, K.; Walton, L.J.; Tunnacliffe, A. LEA proteins prevent protein aggregation due to water stress. Biochem. J. 2005, 388, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Chakrabortee, S.; Tripathi, R.; Watson, M.; Schierle, G.S.K.; Kurniawan, D.P.; Kaminski, C.F.; Wise, M.J.; Tunnacliffe, A. Intrinsically disordered proteins as molecular shields. Mol. Biosyst. 2012, 8, 210–219. [Google Scholar] [CrossRef]
- Hughes, S.L.; Schart, V.; Malcolmson, J.; Hogarth, K.A.; Martynowicz, D.M.; Tralman-Baker, E.; Patel, S.N.; Graether, S.P. The importance of size and disorder in the cryoprotective effects of dehydrins. Plant Physiol. 2013, 163, 1376–1386. [Google Scholar] [CrossRef]
- Palmer, S.R.; De Villa, R.; Graether, S.P. Sequence composition versus sequence order in the cryoprotective function of an intrinsically disordered stress-response protein. Protein Sci. 2019, 28, 1448–1459. [Google Scholar] [CrossRef]
- Smith, M.A.; Graether, S.P. The disordered dehydrin and its role in plant protection: A biochemical perspective. Biomolecules 2022, 12, 294. [Google Scholar] [CrossRef]
- Ferreira, L.A.; Walczyk Mooradally, A.; Zaslavsky, B.; Uversky, V.N.; Graether, S.P. Effect of an intrinsically disordered plant stress protein on the properties of water. Biophys. J. 2018, 115, 1696–1706. [Google Scholar]
- Hernández-Sánchez, I.E.; Maruri-López, I.; Martinez-Martinez, C.; Janis, B.; Jiménez-Bremont, J.F.; Covarrubias, A.A.; Menze, M.A.; Graether, S.P.; Thalhammer, A. LEAfing through literature: Late embryogenesis abundant proteins coming of age-achievements and perspectives. J. Exp. Bot. 2022, 73, 6525–6546. [Google Scholar]
- Abdul Aziz, M.; Masmoudi, K. Multifaceted roles of versatile LEA-II proteins in plants. In Multiple Biotic Stress Tolerances in Higher Plants: Addressing the Growing Challenges; CRC Press: Boca Raton, FL, USA, 2023; pp. 143–162. [Google Scholar]
- Matsuo, N.; Goda, N.; Shimizu, K.; Fukuchi, S.; Ota, M.; Hiroaki, H. Discovery of cryoprotective activity in human genome-derived intrinsically disordered proteins. Int. J. Mol. Sci. 2018, 19, 401. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, J.; Bryant, G. Freezing, drying, and/or vitrification of membrane-solute-water systems. Cryobiology 1999, 39, 103–129. [Google Scholar] [CrossRef] [PubMed]
- Egerton-Warburton, L.M.; Balsamo, R.A.; Close, T.J. Temporal accumulation and ultrastructural localization of dehydrins in Zea mays. Physiol. Plant. 1997, 101, 545–555. [Google Scholar] [CrossRef]
- Danyluk, J.; Perron, A.; Houde, M.; Limin, A.; Fowler, B.; Benhamou, N.; Sarhan, F. Accumulation of an acidic dehydrin in the vicinity of the plasma membrane during cold acclimation of wheat. Plant Cell 1998, 10, 623–638. [Google Scholar] [CrossRef] [PubMed]
- Puhakainen, T.; Hess, M.W.; Mäkelä, P.; Svensson, J.; Heino, P.; Palva, E.T. Overexpression of multiple dehydrin genes enhances tolerance to freezing stress in Arabidopsis. Plant Mol. Biol. 2004, 54, 743–753. [Google Scholar] [CrossRef] [PubMed]
- Szabala, B.M.; Fudali, S.; Rorat, T. Accumulation of acidic SK3 dehydrins in phloem cells of cold- and drought-stressed plants of the Solanaceae. Planta 2014, 239, 847–863. [Google Scholar] [CrossRef]
- Murray, M.R.; Graether, S.P. Physiological, structural, and functional insights into the cryoprotection of membranes by the dehydrins. Front. Plant Sci. 2022, 13, 886525. [Google Scholar]
- Hara, M.; Terashima, S.; Fukaya, T.; Kuboi, T. Enhancement of cold tolerance and inhibition of lipid peroxidation by citrus dehydrin in transgenic tobacco. Planta 2003, 217, 290–298. [Google Scholar]
- Hara, M.; Fujinaga, M.; Kuboi, T. Radical scavenging activity and oxidative modification of citrus dehydrin. Plant Physiol. Biochem. 2004, 42, 657–662. [Google Scholar] [CrossRef]
- Hara, M.; Fujinaga, M.; Kuboi, T. Metal binding by citrus dehydrin with histidine-rich domains. J. Exp. Bot. 2005, 56, 2695–2703. [Google Scholar] [CrossRef]
- Sun, X.; Lin, H.H. Role of plant dehydrins in antioxidation mechanisms. Biologia 2010, 65, 755–759. [Google Scholar]
- Hodges, D.M.; DeLong, J.M.; Forney, C.F.; Prange, R.K. Improving the thiobarbituric acid-reactive-substances assay for estimating lipid peroxidation in plant tissues containing anthocyanin and other interfering compounds. Planta 1999, 207, 604–611. [Google Scholar]
- Xu, H.; Yang, Y.; Xie, L.; Li, X.; Feng, C.; Chen, J.; Xu, C. Involvement of multiple types of dehydrins in the freezing response in loquat (Eriobotrya japonica). PLoS ONE 2014, 9, e87575. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Li, P.; Li, C.; Pan, Y.; Jiang, X.; Zhu, D.; Zhao, Q.; Yu, J. SiLEA14, a novel atypical LEA protein, confers abiotic stress resistance in foxtail millet. BMC Plant Biol. 2014, 14, 290. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Wang, X.; Zhang, L. Structural and functional dynamics of dehydrins: A plant protector protein under abiotic stress. Int. J. Mol. Sci. 2018, 19, 3420. [Google Scholar] [CrossRef]
- Atkinson, J.; Clarke, M.W.; Warnica, J.M.; Boddington, K.F.; Graether, S.P. Structure of an intrinsically disordered stress protein alone and bound to a membrane surface. Biophys. J. 2016, 111, 480–491. [Google Scholar] [CrossRef] [PubMed]
- Eriksson, S.K.; Eremina, N.; Barth, A.; Danielsson, J.; Harryson, P. Membrane-induced folding of the plant stress dehydrin Lti30. Plant Physiol. 2016, 171, 932–943. [Google Scholar]
- Koag, M.C.; Wilkens, S.; Fenton, R.D.; Resnik, J.; Vo, E.; Close, T.J. The K-segment of maize DHN1 mediates binding to anionic phospholipid vesicles and concomitant structural changes. Plant Physiol. 2009, 150, 1503–1514. [Google Scholar]
- Andersson, J.M.; Pham, Q.D.; Mateos, H.; Eriksson, S.; Harryson, P.; Sparr, E. The plant dehydrin Lti30 stabilizes lipid lamellar structures in varying hydration conditions. J. Lipid Res. 2020, 61, 1014–1024. [Google Scholar] [PubMed]
- Rahman, L.N.; Chen, L.; Nazim, S.; Bamm, V.V.; Yaish, M.W.; Moffatt, B.A.; Dutcher, J.R.; Harauz, G. Interactions of intrinsically disordered Thellungiella salsuginea dehydrins TsDHN-1 and TsDHN-2 with membranes—Synergistic effects of lipid composition and temperature on secondary structure. Biochem. Cell Biol. 2010, 88, 791–807. [Google Scholar] [PubMed]
- Rahman, L.N.; McKay, F.; Giuliani, M.; Quirk, A.; Moffatt, B.A.; Harauz, G.; Dutcher, J.R. Interactions of Thellungiella salsuginea dehydrins TsDHN-1 and TsDHN-2 with membranes at cold and ambient temperatures—Surface morphology and single-molecule force measurements show phase separation, and reveal tertiary and quaternary associations. Biochim. Biophys. Acta 2013, 1828, 967–980. [Google Scholar] [PubMed]
- Goldgur, Y.; Rom, S.; Ghirlando, R.; Shkolnik, D.; Shadrin, N.; Konrad, Z.; Bar-Zvi, D. Desiccation and zinc binding induce transition of tomato Abscisic Acid Stress Ripening 1, a water stress- and salt stress-regulated plant-specific protein, from unfolded to folded state. Plant Physiol. 2007, 143, 617–628. [Google Scholar] [PubMed]
- Fonin, A.V.; Darling, A.L.; Kuznetsova, I.M.; Turoverov, K.K.; Uversky, V.N. Intrinsically disordered proteins in crowded milieu: When chaos prevails within the cellular gumbo. Cell. Mol. Life Sci. 2018, 75, 3907–3929. [Google Scholar] [CrossRef]
- Tulumello, D.V.; Deber, C.M. SDS micelles as a membrane-mimetic environment for transmembrane segments. Biochemistry 2009, 48, 12096–12103. [Google Scholar] [CrossRef]
- Ghanmi, S.; Smith, M.A.; Zaidi, I.; Drira, M.; Graether, S.P.; Hanin, M. Isolation and molecular characterization of an FSK2-type dehydrin from Atriplex halimus. Phytochemistry 2023, 213, 113783. [Google Scholar]
- Tolleter, D.; Jaquinod, M.; Mangavel, C.; Passirani, C.; Saulnier, P.; Manon, S.; Teyssier, E.; Payet, N.; Avelange-Macherel, M.H.; Macherel, D. Structure and function of a mitochondrial late embryogenesis abundant protein are revealed by desiccation. Plant Cell 2007, 19, 1580–1589. [Google Scholar]
- Shih, M.D.; Hsieh, T.Y.; Lin, T.P.; Hsing, Y.I.; Hoekstra, F.A. Characterization of two soybean (Glycine max L.) LEA IV proteins by circular dichroism and Fourier transform infrared spectrometry. Plant Cell Physiol. 2010, 51, 395–407. [Google Scholar] [CrossRef]
- Shih, M.D.; Hsieh, T.Y.; Jian, W.T.; Wu, M.T.; Yang, S.J.; Hoekstra, F.A.; Hsing, Y.I.C. Functional studies of soybean (Glycine max L.) seed LEA proteins GmPM6, GmPM11, and GmPM30 by CD and FTIR spectroscopy. Plant Sci. 2012, 196, 152–159. [Google Scholar] [CrossRef]
- Peter, B.; Ngubane, N.C.; Fanucchi, S.; Dirr, H.W. Membrane mimetics induce helix formation and oligomerization of the chloride intracellular channel protein 1 transmembrane domain. Biochemistry 2013, 52, 2739–2749. [Google Scholar]
- Vargas, C.; Arenas, R.C.; Frotscher, E.; Keller, S. Nanoparticle self-assembly in mixtures of phospholipids with styrene/maleic acid copolymers or fluorinated surfactants. Nanoscale 2015, 7, 20685–20696. [Google Scholar] [CrossRef] [PubMed]
- Popova, A.V.; Hundertmark, M.; Seckler, R.; Hincha, D.K. Structural transitions in the intrinsically disordered plant dehydration stress protein LEA7 upon drying are modulated by the presence of membranes. Biochim. Biophys. Acta 2011, 1808, 1879–1887. [Google Scholar] [CrossRef]
- Thalhammer, A.; Hundertmark, M.; Popova, A.V.; Seckler, R.; Hincha, D.K. Interaction of two intrinsically disordered plant stress proteins (COR15A and COR15B) with lipid membranes in the dry state. Biochim. Biophys. Acta 2010, 1798, 1812–1820. [Google Scholar] [CrossRef]
- Thalhammer, A.; Bryant, G.; Sulpice, R.; Hincha, D.K. Disordered cold regulated 15 proteins protect chloroplast membranes during freezing through binding and folding, but do not stabilize chloroplast enzymes in vivo. Plant Physiol. 2014, 166, 190–201. [Google Scholar] [CrossRef]
- Bremer, A.; Kent, B.; Hauß, T.; Thalhammer, A.; Yepuri, N.R.; Darwish, T.A.; Garvey, C.J.; Bryant, G.; Hincha, D.K. Intrinsically disordered stress protein COR15A resides at the membrane surface during dehydration. Biophys. J. 2017, 113, 572–579. [Google Scholar] [CrossRef] [PubMed]
- Shou, K.; Bremer, A.; Rindfleisch, T.; Knox-Brown, P.; Hirai, M.; Rekas, A.; Garvey, C.J.; Hincha, D.K.; Stadler, A.M.; Thalhammer, A. Conformational selection of the intrinsically disordered plant stress protein COR15A in response to solution osmolarity—An X-ray and light scattering study. Phys. Chem. Chem. Phys. 2019, 21, 18727–18740. [Google Scholar] [CrossRef]
- Meng, Y.C.; Zhang, H.F.; Pan, X.X.; Chen, N.; Hu, H.F.; Haq, S.U.; Khan, A.; Chen, R.G. CaDHN3, a pepper (Capsicum annuum L.) dehydrin gene enhances the tolerance against salt and drought stresses by reducing ROS accumulation. Int. J. Mol. Sci. 2021, 22, 3205. [Google Scholar] [PubMed]
- Hernández-Sánchez, I.E.; Maruri-López, I.; Graether, S.P.; Jiménez-Bremont, J.F. In vivo evidence for homo- and heterodimeric interactions of Arabidopsis thaliana dehydrins AtCOR47, AtERD10, and AtRAB18. Sci. Rep. 2017, 7, 17036. [Google Scholar] [CrossRef]
- Upadhyaya, G.; Das, A.; Basu, C.; Agarwal, T.; Basak, C.; Chakraborty, C.; Halder, T.; Basu, G.; Ray, S. Multiple copies of a novel amphipathic α-helix forming segment in Physcomitrella patens dehydrin play a key role in abiotic stress mitigation. J. Biol. Chem. 2021, 296, 100596. [Google Scholar] [CrossRef]
- Hernández-Sánchez, I.E.; Maruri-López, I.; Molphe-Balch, E.P.; Becerra-Flora, A.; Jaimes-Miranda, F.; Jiménez-Bremont, J.F. Evidence for in vivo interactions between dehydrins and the aquaporin AtPIP2B. Biochem. Biophys. Res. Commun. 2019, 510, 545–550. [Google Scholar] [CrossRef] [PubMed]
- Hasan, M.K.; Cheng, Y.; Kanwar, M.K.; Chu, X.Y.; Ahammed, G.J.; Qi, Z.Y. Responses of plant proteins to heavy metal stress-A review. Front. Plant Sci. 2017, 8, 1492. [Google Scholar] [PubMed]
- Zhang, Y.; Wang, Y.; Wen, W.; Shi, Z.; Gu, Q.; Ahammed, G.J.; Cao, K.; Shah Jahan, M.; Shu, S.; Wang, J.; et al. Hydrogen peroxide mediates spermidine-induced autophagy to alleviate salt stress in cucumber. Autophagy 2021, 17, 2876–2890. [Google Scholar] [PubMed]
- Li, X.; Liu, Q.; Feng, H.; Deng, J.; Zhang, R.; Wen, J.; Dong, J.; Wang, T. Dehydrin MtCAS31 promotes autophagic degradation under drought stress. Autophagy 2020, 16, 862–877. [Google Scholar] [CrossRef] [PubMed]
- Lv, A.; Wen, W.; Fan, N.; Su, L.; Zhou, P.; An, Y. Dehydrin MsDHN1 improves aluminum tolerance of alfalfa (Medicago sativa L.) by affecting oxalate exudation from root tips. Plant J. 2021, 108, 441–458. [Google Scholar] [CrossRef]
- Raffaele, S.; Mongrand, S.; Gamas, P.; Niebel, A.; Ott, T. Genome-wide annotation of remorins, a plant-specific protein family: Evolutionary and functional perspectives. Plant Physiol. 2007, 145, 593–600. [Google Scholar] [CrossRef] [PubMed]
- Gouguet, P.; Gronnier, J.; Legrand, A.; Perraki, A.; Jolivet, M.D.; Deroubaix, A.F.; German-Retana, S.; Boudsocq, M.; Habenstein, B.; Mongrand, S.; et al. Connecting the dots: From nanodomains to physiological functions of REMORINs. Plant Physiol. 2021, 185, 632–649. [Google Scholar] [PubMed]
- Gronnier, J.; Crowet, J.M.; Habenstein, B.; Nasir, M.N.; Bayle, V.; Hosy, E.; Platre, M.P.; Gouguet, P.; Raffaele, S.; Martinez, D.; et al. Structural basis for plant plasma membrane protein dynamics and organization into functional nanodomains. Elife 2017, 6, 26404. [Google Scholar] [CrossRef]
- Liang, P.; Stratil, T.F.; Popp, C.; Marı’n, M.; Folgmann, J.; Mysore, K.S.; Wen, J.; Ott, T. Symbiotic root infections in Medicago truncatula require remorin-mediated receptor stabilization in membrane nanodomains. Proc. Natl. Acad. Sci. USA 2018, 115, 5289–5294. [Google Scholar] [CrossRef]
- Kohorn, B.D.; Hoon, D.; Minkoff, B.B.; Sussman, M.R.; Kohorn, S.L. Rapid oligo-galacturonide induced changes in protein phosphorylation in Arabidopsis. Mol. Cell. Proteom. 2016, 15, 1351–1359. [Google Scholar]
- Menz, J.; Li, Z.; Schulze, W.X.; Ludewig, U. Early nitrogen-deprivation responses in Arabidopsis roots reveal distinct differences on transcriptome and (phospho-) proteome levels between nitrate and ammonium nutrition. Plant J. 2016, 88, 717–734. [Google Scholar] [CrossRef]
- Nikonorova, N.; Van Den Broeck, L.; Zhu, S.; Van De Cotte, B.; Dubois, M.; Gevaert, K.; Inzé, D.; De Smet, I. Early mannitol-triggered changes in the Arabidopsis leaf (phospho)proteome reveal growth regulators. J. Exp. Bot. 2018, 69, 4591–4607. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Hsu, C.C.; Du, Y.; Zhu, P.; Zhao, C.; Fu, X.; Zhang, C.; Paez, J.S.; Macho, A.P.; Tao, W.A.; et al. Mapping proteome-wide targets of protein kinases in plant stress responses. Proc. Natl. Acad. Sci. USA 2020, 117, 3270–3280. [Google Scholar] [PubMed]
- Raffaele, S.; Bayer, E.; Lafarge, D.; Cluzet, S.; German Retana, S.; Boubekeur, T.; Leborgne-Castel, N.; Carde, J.-P.; Lherminier, J.; Noirot, E.; et al. Remorin, a solanaceae protein resident in membrane rafts and plasmodesmata, impairs potato virus X movement. Plant Cell 2009, 21, 1541–1555. [Google Scholar] [CrossRef] [PubMed]
- Perraki, A.; Cacas, J.L.; Crowet, J.M.; Lins, L.; Castroviejo, M.; German-Retana, S.; Mongrand, S.; Raffaele, S. Plasma membrane localization of Solanum tuberosum remorin from group 1, homolog 3 is mediated by conformational changes in a novel C-terminal anchor and required for the restriction of potato virus X movement. Plant Physiol. 2012, 160, 624–637. [Google Scholar] [CrossRef] [PubMed]
- Perraki, A.; Gronnier, J.; Gouguet, P.; Boudsocq, M.; Deroubaix, F.; Simon, V.; Legrand, A.; German-Retana, S.; Zipfel, C.; Bayer, E.M.F.; et al. REM1.3′s phospho-status defines its plasma membrane nanodomain organization and activity in restricting PVX cell-to-cell movement. PLoS Pathog. 2018, 14, e1007378. [Google Scholar] [CrossRef] [PubMed]
- Checker, V.G.; Khurana, P. Molecular and functional characterization of mulberry EST encoding remorin (MiREM) involved in abiotic stress. Plant Cell Rep. 2013, 32, 1729–1741. [Google Scholar] [CrossRef] [PubMed]
- Yue, J.; Li, C.; Liu, Y.; Yu, J. A remorin gene SiREM6, the target gene of SiARDP, from foxtail millet (Setaria italica) promotes high salt tolerance in transgenic Arabidopsis. PLoS ONE 2014, 9, e100772. [Google Scholar] [CrossRef]
- Zhang, H.; Deng, C.; Wu, X.; Yao, J.; Zhang, Y.; Zhang, Y.; Deng, S.; Zhao, N.; Zhao, R.; Zhou, X.; et al. Populus euphratica remorin 6.5 activates plasma membrane H + -ATPases to mediate salt tolerance. Tree Physiol. 2020, 40, 731–745. [Google Scholar] [CrossRef]
- Jarsch, I.K.; Ott, T. Perspectives on remorin proteins, membrane rafts, and their role during plant-microbe interactions. Mol. Plant Microbe Interact. 2011, 24, 7–12. [Google Scholar]
- Liu, G.; Xu, H.; Zhang, L.; Zheng, Y. Fe binding properties of two soybean (Glycine max L.) LEA4 proteins associated with antioxidant activity. Plant Cell Physiol. 2011, 52, 994–1002. [Google Scholar] [CrossRef]
- Graether, S.P.; Boddington, K.F. Disorder and function: A review of the dehydrin protein family. Front. Plant Sci. 2014, 5, 576. [Google Scholar] [CrossRef]
- French-Pacheco, L.; Cuevas-Velazquez, C.L.; Rivillas-Acevedo, L.; Covarrubias, A.A.; Amero, C. Metal-binding polymorphism in late embryogenesis abundant protein AtLEA4-5, an intrinsically disordered protein. PeerJ 2018, 6, e4930. [Google Scholar] [CrossRef] [PubMed]
- Golan, I.; Dominguez, P.G.; Konrad, Z.; Shkolnik-Inbar, D.; Carrari, F.; Bar-Zvi, D. Tomato Abscisic Acid Stress Ripening (ASR) gene family revisited. PLoS ONE 2014, 9, e107117. [Google Scholar]
- Philippe, R.; Courtois, B.; Mcnally, K.L.; Mournet, P.; El-Malki, R.; Le Paslier, M.C.; Fabre, D.; Billot, C.; Brunel, D.; Glaszmann, J.C.; et al. Structure, allelic diversity and selection of Asr genes, candidate for drought tolerance, in Oryza sativa L. and wild relatives. Theor. Appl. Genet. 2010, 121, 769–787. [Google Scholar] [CrossRef]
- Li, H.; Guan, H.; Zhuo, Q.; Wang, Z.; Li, S.; Si, J.; Zhang, B.; Feng, B.; Kong, L.A.; Wang, F.; et al. Genome-wide characterization of the abscisic acid-, stress- and ripening-induced (ASR) gene family in wheat (Triticum aestivum L.). Biol. Res. 2020, 53, 23. [Google Scholar]
- Hong, S.H.; Kim, I.J.; Yang, D.C.; Chung, W.I. Characterization of an abscisic acid responsive gene homologue from Cucumis melo. J. Exp. Bot. 2002, 53, 2271–2272. [Google Scholar]
- Jeanneau, M.; Gerentes, D.; Foueillassar, X.; Zivy, M.; Vidal, J.; Toppan, A.; Perez, P. Improvement of drought tolerance in maize: Towards the functional validation of the Zm-Asr1 gene and increase of water use efficiency by over-expressing C4-PEPC. Biochimie 2002, 84, 1127–1135. [Google Scholar] [CrossRef] [PubMed]
- Virlouvet, L.; Jacquemot, M.P.; Gerentes, D.; Corti, H.; Bouton, S.; Gilard, F.; Valot, B.; Trouverie, J.; Tcherkez, G.; Falque, M.; et al. The ZmASR1 protein influences branched-chain amino acid biosynthesis and maintains kernel yield in maize under water-limited conditions. Plant Physiol. 2011, 157, 917–936. [Google Scholar] [CrossRef]
- Liu, H.Y.; Dai, J.R.; Feng, D.R.; Liu, B.; Wang, H.B.; Wang, J.F. Characterization of a novel plantain Asr gene, MpAsr, that is regulated in response to infection of Fusarium oxysporum f. sp. cubense and abiotic stresses. J. Integr. Plant Biol. 2010, 52, 315–323. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Huang, C.; Deng, X.; Zhou, S.; Chen, L.; Li, Y.; Wang, C.; Ma, Z.; Yuan, Q.; Wang, Y.; et al. TaASR1, a transcription factor gene in wheat, confers drought stress tolerance in transgenic tobacco. Plant Cell Environ. 2013, 36, 1449–1464. [Google Scholar] [CrossRef]
- Li, J.; Li, Y.; Yin, Z.; Jiang, J.; Zhang, M.; Guo, X.; Ye, Z.; Zhao, Y.; Xiong, H.; Zhang, Z.; et al. OsASR5 enhances drought tolerance through a stomatal closure pathway associated with ABA and H2O2 signalling in rice. Plant Biotechnol. J. 2017, 15, 183–196. [Google Scholar] [CrossRef] [PubMed]
- Park, S.I.; Kim, J.J.; Shin, S.Y.; Kim, Y.S.; Yoon, H.S. ASR enhances environmental stress tolerance and improves grain yield by modulating stomatal closure in rice. Front. Plant Sci. 2020, 10, 1752. [Google Scholar] [CrossRef]
- Cakir, B.; Agasse, A.; Gaillard, C.; Saumonneau, A.; Delrot, S.; Atanassova, R. A grape ASR protein involved in sugar and abscisic acid signaling. Plant Cell 2003, 15, 2165–2180. [Google Scholar] [PubMed]
- Jia, H.; Jiu, S.; Zhang, C.; Wang, C.; Tariq, P.; Liu, Z.; Wang, B.; Cui, L.; Fang, J. Abscisic acid and sucrose regulate tomato and strawberry fruit ripening through the abscisic acid-stress-ripening transcription factor. Plant Biotechnol. J. 2016, 14, 2045–2065. [Google Scholar] [CrossRef] [PubMed]
- Kalifa, Y.; Pearlson, E.; Gilad, A.; Konrad, Z.; Scolnik, P.A.; Bar-Zvi, D. Overexpression of the water and salt stress-regulated Asr1 gene confers an increased salt tolerance. Plant Cell Environ. 2004, 27, 1459–1468. [Google Scholar] [CrossRef]
- Yang, C.Y.; Chen, Y.C.; Jauh, G.Y.; Wang, C.S. A Lily ASR protein involves abscisic acid signaling and confers drought and salt resistance in Arabidopsis. Plant Physiol. 2005, 139, 836–846. [Google Scholar] [CrossRef]
- Hu, Y.X.; Yang, X.; Li, X.L.; Yu, X.D.; Li, Q.L. The SlASR gene cloned from the extreme halophyte Suaeda liaotungensis K. enhances abiotic stress tolerance in transgenic Arabidopsis thaliana. Gene 2014, 549, 243–251. [Google Scholar] [CrossRef]
- Kalifa, Y.; Gilad, A.; Konrad, Z.; Zaccai, M.; Scolnik, P.A.; Bar-Zvi, D. The water- and salt-stress regulated Asr1 gene encodes a zinc-dependent DNA-binding protein. Biochem. J. 2004, 381, 373–378. [Google Scholar]
- Ricardi, M.M.; González, R.M.; Zhong, S.; Domínguez, P.G.; Duffy, T.; Turjanski, P.G.; Salgado Salter, J.D.; Alleva, K.; Carrari, F.; Giovannoni, J.J.; et al. Genome-wide data (ChIP-seq) enabled identification of cell wall-related and aquaporin genes as targets of tomato ASR1, a drought stress-responsive transcription factor. BMC Plant Biol. 2014, 14, 29. [Google Scholar] [CrossRef]
- Rom, S.; Gilad, A.; Kalifa, Y.; Konrad, Z.; Karpasas, M.M.; Goldgur, Y.; Bar-Zvi, D. Mapping the DNA- and zinc-binding domains of ASR1 (abscisic acid stress ripening), an abiotic-stress regulated plant specific protein. Biochimie 2006, 88, 621–628. [Google Scholar] [CrossRef] [PubMed]
- Wetzler, D.E.; Fuchs Wightman, F.; Bucci, H.A.; Rinaldi, J.; Caramelo, J.J.; Iusem, N.D.; Ricardi, M.M. Conformational plasticity of the intrinsically disordered protein ASR1 modulates its function as a drought stress-responsive gene. PLoS ONE 2018, 13, e0202808. [Google Scholar] [CrossRef] [PubMed]
- Hamdi, K.; Salladini, E.; O’Brien, D.P.; Brier, S.; Chenal, A.; Yacoubi, I.; Longhi, S. Structural disorder and induced folding within two cereal, ABA stress and ripening (ASR) proteins. Sci. Rep. 2017, 7, 15544. [Google Scholar] [CrossRef]
- Barros, N.L.F.; Siqueira, A.S.; Arenhart, R.A.; Margis-Pinheiro, M. Modeling the zinc effect on OsASR5-STAR1 promoter interaction by molecular dynamics. Proteins 2023, 91, 944–955. [Google Scholar] [CrossRef]
- Laity, J.H.; Lee, B.M.; Wright, P.E. Zinc finger proteins: New insights into structural and functional diversity. Curr. Opin. Struct. Biol. 2001, 11, 39–46. [Google Scholar] [CrossRef]
- Yacoubi, I.; Hamdi, K.; Fourquet, P.; Bignon, C.; Longhi, S. Structural and functional characterization of the ABA-water deficit stress domain from wheat and barley: An intrinsically disordered domain behind the versatile functions of the Plant abscissic acid, stress and ripening protein family. Int. J. Mol. Sci. 2021, 22, 2314. [Google Scholar] [PubMed]
- Liu, Y.; Zhang, H.; Han, J.; Jiang, S.; Geng, X.; Xue, D.; Chen, Y.; Zhang, C.; Zhou, Z.; Zhang, W.; et al. Functional assessment of hydrophilic domains of late embryogenesis abundant proteins from distant organisms. Microb. Biotechnol. 2019, 12, 752–762. [Google Scholar] [CrossRef]
- Murvai, N.; Kalmar, L.; Szalaine Agoston, B.; Szabo, B.; Tantos, A.; Csikos, G.; Micsonai, A.; Kardos, J.; Vertommen, D.; Nguyen, P.N.; et al. Interplay of structural disorder and short binding elements in the cellular chaperone function of plant dehydrin ERD14. Cells 2020, 9, 1856. [Google Scholar] [CrossRef]
- Murvai, N.; Kalmar, L.; Szabo, B.; Schad, E.; Micsonai, A.; Kardos, J.; Buday, L.; Han, K.H.; Tompa, P.; Tantos, A. Cellular chaperone function of intrinsically disordered dehydrin ERD14. Int. J. Mol. Sci. 2021, 22, 6190. [Google Scholar]
- Dang, N.X.; Popova, A.V.; Hundertmark, M.; Hincha, D.K. Functional characterization of selected LEA proteins from Arabidopsis thaliana in yeast and in vitro. Planta 2014, 240, 325–336. [Google Scholar]
- Saucedo, A.L.; Hernández-Domínguez, E.E.; de Luna-Valdez, L.A.; Guevara-García, A.A.; Escobedo-Moratilla, A.; Bojorquéz-Velázquez, E.; Del Río-Portilla, F.; Fernández-Velasco, D.A.; Barba de la Rosa, A.P. Insights on structure and function of a late embryogenesis abundant protein from Amaranthus cruentus: An intrinsically disordered protein Involved in protection against desiccation, oxidant conditions, and osmotic stress. Front. Plant Sci. 2017, 8, 497. [Google Scholar] [CrossRef]
- Gandass, N.; Kajal; Salvi, P. Intrinsically disordered protein, DNA binding with one finger transcription factor (OsDOF27) implicates thermotolerance in yeast and rice. Front. Plant Sci. 2022, 13, 956299. [Google Scholar]
- Mahjoubi, H.; Ebel, C.; Hanin, M. Molecular and functional characterization of the durum wheat TdRL1, a member of the conserved Poaceae RSS1-like family that exhibits features of intrinsically disordered proteins and confers stress tolerance in yeast. Funct. Integr. Genom. 2015, 15, 717–728. [Google Scholar] [CrossRef]
- Mahjoubi, H.; Tamari, Y.; Takeda, S.; Bouchabké-Coussa, O.; Hanin, M.; Herzog, E.; Schmit, A.C.; Chabouté, M.E.; Ebel, C. The wheat TdRL1 is the functional homolog of the rice RSS1 and promotes plant salt stress tolerance. Plant Cell Rep. 2018, 37, 1625–1637. [Google Scholar] [CrossRef] [PubMed]
- Arakawa, K. Examples of extreme survival: Tardigrade genomics and molecular anhydrobiology. Annu. Rev. Anim. Biosci. 2022, 10, 17–37. [Google Scholar] [CrossRef] [PubMed]
- Majumdar, A.; Dogra, P.; Maity, S.; Mukhopadhyay, S. Liquid–liquid phase separation is driven by large-scale conformational unwinding and fluctuations of intrinsically disordered protein molecules. J. Phys. Chem. Lett. 2019, 10, 3929–3936. [Google Scholar] [CrossRef] [PubMed]
- Fang, X.; Wang, L.; Ishikawa, R.; Li, Y.; Fiedler, M.; Liu, F.; Calder, G.; Rowan, B.; Weigel, D.; Li, P.; et al. Arabidopsis FLL2 promotes liquid-liquid phase separation of polyadenylation complexes. Nature 2019, 569, 265–269. [Google Scholar]
- Gutierrez-Beltran, E.; Elander, P.H.; Dalman, K.; Dayhoff II, G.W.; Moschou, P.N.; Uversky, V.N.; Crespo, J.L.; Bozhkov, P.V. Tudor staphylococcal nuclease is a docking platform for stress granule components and is essential for SnRK1 activation in Arabidopsis. EMBO J. 2021, 40, e105043. [Google Scholar] [CrossRef]
- Maruri-López, I.; Figueroa, N.E.; Hernández-Sánchez, I.E.; Chodasiewicz, M. Plant stress granules: Trends and beyond. Front. Plant Sci. 2021, 12, 722643. [Google Scholar] [CrossRef]
- Jung, J.H.; Barbosa, A.D.; Hutin, S.; Kumita, J.R.; Gao, M.; Derwort, D.; Silva, C.S.; Lai, X.; Pierre, E.; Geng, F.; et al. A prion-like domain in ELF3 functions as a thermosensor in Arabidopsis. Nature 2020, 585, 256–260. [Google Scholar] [CrossRef]
- Zhu, S.; Gu, J.; Yao, J.; Li, Y.; Zhang, Z.; Xia, W.; Wang, Z.; Gui, X.; Li, L.; Li, D.; et al. Liquid–liquid phase separation of RBGD2/4 is required for heat stress resistance in Arabidopsis. Dev. Cell 2022, 57, 583–597. [Google Scholar] [CrossRef]
- Zhu, P.; Lister, C.; Dean, C. Cold-induced Arabidopsis FRIGIDA nuclear condensates for FLC repression. Nature 2021, 599, 657–661. [Google Scholar]
- Cao, X.; Du, Q.; Guo, Y.; Wang, Y.; Jiao, Y. Condensation of STM is critical for shoot meristem maintenance and salt tolerance in Arabidopsis. Mol. Plant 2023, 16, 1445–1459. [Google Scholar]
- Huang, X.; Chen, S.; Li, W.; Tang, L.; Zhang, Y.; Yang, N.; Zou, Y.; Zhai, X.; Xiao, N.; Liu, W.; et al. ROS regulated reversible protein phase separation synchronizes plant flowering. Nat. Chem. Biol. 2021, 17, 549–557. [Google Scholar] [PubMed]
- Chen, D.; Lyu, M.; Kou, X.; Li, J.; Yang, Z.; Gao, L.; Li, Y.; Fan, L.M.; Shi, H.; Zhong, S. Integration of light and temperature sensing by liquid-liquid phase separation of phytochrome B. Mol. Cell 2022, 82, 3015–3029. [Google Scholar] [CrossRef]
- Dorone, Y.; Boeynaems, S.; Flores, E.; Jin, B.; Hateley, S.; Bossi, F. A prion-like protein regulator of seed germination undergoes hydration-dependent phase separation. Cell 2021, 184, 4284–4298. [Google Scholar]
- Wang, B.; Zhang, H.; Huai, J.; Peng, F.; Wu, J.; Lin, R.; Fang, X. Condensation of SEUSS promotes hyperosmotic stress tolerance in Arabidopsis. Nat. Chem. Biol. 2022, 18, 1361–1369. [Google Scholar]
- Marín, M.; Ott, T. Intrinsic disorder in plant proteins and phytopathogenic bacterial effectors. Chem. Rev. 2014, 114, 6912–6932. [Google Scholar] [PubMed]
- Marín, M.; Uversky, V.N.; Ott, T. Intrinsic disorder in pathogen effectors: Protein flexibility as an evolutionary hallmark in a molecular arms race. Plant Cell 2013, 25, 3153–3157. [Google Scholar] [CrossRef] [PubMed]
- Molisso, D.; Coppola, M.; Buonanno, M.; Di Lelio, I.; Aprile, A.M.; Langella, E.; Rigano, M.M.; Francesca, S.; Chiaiese, P.; Palmieri, G.; et al. Not only systemin: Prosystemin harbors other active regions able to protect tomato plants. Front. Plant Sci. 2022, 13, 887674. [Google Scholar]
- Belkhadir, Y.; Nimchuk, Z.; Hubert, D.A.; Mackey, D.; Dangl, J.L. Arabidopsis RIN4 negatively regulates disease resistance mediated by RPS2 and RPM1 downstream or independent of the NDR1 signal modulator and is not required for the virulence functions of bacterial type III effectors AvrRpt2 or AvrRpm1. Plant Cell 2004, 16, 2822–2835. [Google Scholar] [CrossRef]
- Kim, M.G.; da Cunha, L.; McFall, A.J.; Belkhadir, Y.; DebRoy, S.; Dangl, J.L.; Mackey, D. Two Pseudomonas syringae type III effectors inhibit RIN4- regulated basal defense in Arabidopsis. Cell 2005, 121, 749–759. [Google Scholar] [CrossRef]
- Luo, Y.; Caldwell, K.S.; Wroblewski, T.; Wright, M.E.; Michelmore, R.W. Proteolysis of a negative regulator of innate immunity is dependent on resistance genes in tomato and Nicotiana benthamiana and induced by multiple bacterial effectors. Plant Cell 2009, 21, 2458–2472. [Google Scholar] [CrossRef] [PubMed]
- Desveaux, D.; Singer, A.U.; Wu, A.J.; McNulty, B.C.; Musselwhite, L.; Nimchuk, Z.; Sondek, J.; Dangl, J.L. Type III effector activation via nucleotide binding, phosphorylation, and host target interaction. PLoS Pathog. 2007, 3, e48. [Google Scholar]
- Sun, X.; Greenwood, D.R.; Templeton, M.D.; Libich, D.S.; McGhie, T.K.; Xue, B.; Yoon, M.; Cui, W.; Kirk, C.A.; Jones, W.T.; et al. The intrinsically disordered structural platform of the plant defence hub protein RPM1-interacting protein 4 provides insights into its mode of action in the host-pathogen interface and evolution of the nitrate-induced domain protein family. FEBS J. 2014, 281, 3955–3979. [Google Scholar] [CrossRef]
- Toruno, T.Y.; Shen, M.; Coaker, G.; Mackey, D. Regulated disorder: Posttranslational modifications control the RIN4 plant immune signaling hub. Mol. Plant Microbe Interact. 2019, 32, 56–64. [Google Scholar] [CrossRef]
- Contreras, E.; Martinez, M. Comparative and evolutionary analysis of Arabidopsis RIN4-like/NOI proteins induced by herbivory. PLoS ONE 2022, 17, e0270791. [Google Scholar] [CrossRef]
- Kim, H.; Prokchorchik, M.; Sohn, K.H. Investigation of natural RIN4 variants reveals a motif crucial for function and provides an opportunity to broaden NLR regulation specificity. Plant J. 2022, 110, 58–70. [Google Scholar] [CrossRef] [PubMed]
- Jayaraman, J.; Yoon, M.; Hemara, L.M.; Bohne, D.; Tahir, J.; Chen, R.K.Y.; Brendolise, C.; Rikkerink, E.H.A.; Templeton, M.D. Contrasting effector profiles between bacterial colonisers of kiwifruit reveal redundant roles converging on PTI-suppression and RIN4. New Phytol. 2023, 238, 1605–1619. [Google Scholar] [CrossRef]
- Xu, H.; Ye, M.; Xia, A.; Jiang, H.; Huang, P.; Liu, H.; Hou, R.; Wang, Q.; Li, D.; Xu, J.R.; et al. The Fng3 ING protein regulates H3 acetylation and H4 deacetylation by interacting with two distinct histone-modifying complexes. New Phytol. 2022, 235, 2350–2364. [Google Scholar] [CrossRef]
- Mekonnen, G.; Djaja, N.; Yuan, X.; Myong, S. Advanced imaging techniques for studying protein phase separation in living cells and at single-molecule level. Curr. Opin. Chem. Biol. 2023, 76, 102371. [Google Scholar] [CrossRef] [PubMed]
- Ginsawaeng, O.; Heise, C.; Sangwan, R.; Karcher, D.; Hernández-Sánchez, I.E.; Sampathkumar, A.; Zuther, E. Subcellular localization of seed-expressed LEA_4 proteins reveals liquid-liquid phase separation for LEA9 and for LEA48 homo- and LEA42-LEA48 heterodimers. Biomolecules 2021, 11, 1770. [Google Scholar]
- Belott, C.; Janis, B.; Menze, M.A. Liquid-liquid phase separation promotes animal desiccation tolerance. Proc. Natl. Acad. Sci. USA 2020, 117, 27676–27684. [Google Scholar]
- Janis, B.; Belott, C.; Brockman, T.; Menze, M.A. Functional and conformational plasticity of an animal group 1 LEA protein. Biomolecules 2022, 12, 425. [Google Scholar]
- Degand, H.; Faber, A.M.; Dauchot, N.; Mingeot, D.; Watillon, B.; van Cutsem, P.; Morsomme, P.; Boutry, M. Proteomic analysis of chicory root identifies proteins typically involved in cold acclimation. Proteomics 2009, 9, 2903–2907. [Google Scholar] [CrossRef]
- Ghaffari, M.; Toorchi, M.; Valizadeh, M.; Komatsu, S. Differential response of root proteome to drought stress in drought sensitive and tolerant sunflower inbred lines. Funct. Plant Biol. 2013, 40, 609–617. [Google Scholar] [PubMed]
- Li, G.; Peng, X.; Xuan, H.; Wei, L.; Yang, Y.; Guo, T.; Kang, G. Proteomic analysis of leaves and roots of common wheat (Triticum aestivum L.) under copper-stress conditions. J. Proteome Res. 2013, 12, 4846–4861. [Google Scholar] [CrossRef]
- Kosová, K.; Vítámvás, P.; Urban, M.O.; Klíma, M.; Roy, A.; Prášil, I.T. Biological Networks Underlying Abiotic Stress Tolerance in Temperate Crops--A Proteomic Perspective. Int. J. Mol. Sci. 2015, 16, 20913–20942. [Google Scholar] [CrossRef] [PubMed]
- Rahman, L.N.; Smith, G.S.; Bamm, V.V.; Voyer-Grant, J.A.; Moffatt, B.A.; Dutcher, J.R.; Harauz, G. Phosphorylation of Thellungiella salsuginea dehydrins TsDHN-1 and TsDHN-2 facilitates cation-induced conformational changes and actin assembly. Biochemistry 2011, 50, 9587–9604. [Google Scholar] [CrossRef]
- Kumar, S.; Jeevaraj, T.; Yunus, M.H.; Chakraborty, S.; Chakraborty, N. The plant cytoskeleton takes center stage in abiotic stress responses and resilience. Plant Cell Environ. 2023, 46, 5–22. [Google Scholar] [CrossRef]
- Li, J.; Staiger, C.J. Understanding cytoskeletal dynamics during the plant immune response. Annu. Rev. Phytopathol. 2018, 56, 513–533. [Google Scholar] [CrossRef]
- Wang, J.; Lian, N.; Zhang, Y.; Man, Y.; Chen, L.; Yang, H.; Lin, J.; Jing, Y. The cytoskeleton in plant immunity: Dynamics, regulation, and function. Int. J. Mol. Sci. 2022, 23, 15553. [Google Scholar]
- Cheng, M.L.; Lo, S.F.; Hsiao, A.S.; Hong, Y.F.; Yu, S.M.; Ho, T.D. Ectopic expression of WINDING 1 leads to asymmetrical distribution of auxin and a spiral phenotype in rice. Plant Cell Physiol. 2017, 58, 1494–1506. [Google Scholar] [CrossRef] [PubMed]
- Lo, S.F.; Cheng, M.L.; Hsing, Y.C.; Chen, Y.S.; Lee, K.W.; Hong, Y.F.; Hsiao, Y.; Hsiao, A.S.; Chen, P.J.; Wong, L.I.; et al. Rice Big Grain 1 promotes cell division to enhance organ development, stress tolerance and grain yield. Plant Biotechnol. J. 2020, 18, 1969–1983. [Google Scholar] [CrossRef]
- Hsiao, A.S.; Huang, J.Y. Microtubule regulation in plants: From morphological development to stress adaptation. Biomolecules 2023, 13, 627. [Google Scholar] [CrossRef] [PubMed]
- Guharoy, M.; Szabo, B.; Martos, S.C.; Kosol, S.; Tompa, P. Intrinsic structural disorder in cytoskeletal proteins. Cytoskeleton 2013, 70, 550–571. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, J.P.; Frandsen, K.E.H.; Kesten, C. The role of intrinsic disorder in binding of plant microtubule-associated proteins to the cytoskeleton. Cytoskeleton 2023, 80, 404–436. [Google Scholar]
- Kesten, C.; Wallmann, A.; Schneider, R.; McFarlane, H.E.; Diehl, A.; Khan, G.A.; van Rossum, B.J.; Lampugnani, E.R.; Szymanski, W.G.; Cremer, N. The companion of cellulose synthase 1 confers salt tolerance through a Tau-like mechanism in plants. Nat. Commun. 2019, 10, 857. [Google Scholar] [PubMed]
- Hinrichs, M.H.; Jalal, A.; Brenner, B.; Mandelkow, E.; Kumar, S.; Scholz, T. Tau protein diffuses along the microtubule lattice. J. Biol. Chem. 2012, 287, 38559–38568. [Google Scholar] [CrossRef]
- Gui, J.; Liu, C.; Shen, J.; Li, L. Grain setting defect1, encoding a Remorin protein, affects the grain setting in rice through regulating plasmodesmatal conductance. Plant Physiol. 2014, 166, 1463–1478. [Google Scholar]
- Cheng, G.; Yang, Z.; Zhang, H.; Zhang, J.; Xu, J. Remorin interacting with PCaP1 impairs Turnip mosaic virus intercellular movement but is antagonised by VPg. New Phytol. 2020, 225, 2122–2139. [Google Scholar] [CrossRef]
- Ma, Z.; Sun, Y.; Zhu, X.; Yang, L.; Chen, X.; Miao, Y. Membrane nanodomains modulate formin condensation for actin remodeling in Arabidopsis innate immune responses. Plant Cell 2022, 34, 374–394. [Google Scholar] [CrossRef]
- King, M.R.; Petry, S. Phase separation of TPX2 enhances and spatially coordinates microtubule nucleation. Nat. Commun. 2020, 11, 270. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Zhu, X.; Li, C.; Ma, Z.; Han, X.; Luo, Y.; Yang, L.; Yu, J.; Miao, Y. Xanthomonas effector XopR hijacks host actin cytoskeleton via complex coacervation. Nat. Commun. 2021, 12, 4064. [Google Scholar] [CrossRef]
- Pietrosemoli, N.; García-Martín, J.A.; Solano, R.; Pazos, F. Genome-wide analysis of protein disorder in Arabidopsis thaliana: Implications for plant environmental adaptation. PLoS ONE 2013, 8, e55524. [Google Scholar] [CrossRef] [PubMed]
- Camacho-Zarco, A.R.; Schnapka, V.; Guseva, S.; Abyzov, A.; Adamski, W.; Milles, S.; Jensen, M.R.; Zidek, L.; Salvi, N.; Blackledge, M. NMR provides unique insight into the functional dynamics and interactions of intrinsically disordered proteins. Chem. Rev. 2022, 122, 9331–9356. [Google Scholar] [CrossRef] [PubMed]
- Ando, T. Functional implications of dynamic structures of intrinsically disordered proteins revealed by high-speed AFM Imaging. Biomolecules 2022, 12, 1876. [Google Scholar] [CrossRef]
- Chowdhury, A.; Nettels, D.; Schuler, B. Interaction dynamics of intrinsically disordered proteins from single-molecule spectroscopy. Annu. Rev. Biophys. 2023, 52, 433–462. [Google Scholar] [CrossRef]
- LeBlanc, S.J.; Kulkarni, P.; Weninger, K.R. Single molecule FRET: A powerful tool to study intrinsically disordered proteins. Biomolecules 2018, 8, 140. [Google Scholar]
- Metskas, L.A.; Rhoades, E. Single-molecule FRET of intrinsically disordered proteins. Annu. Rev. Phys. Chem. 2020, 71, 391–414. [Google Scholar]
- Hsiao, A.S.; Huang, J.Y. Bioimaging tools move plant physiology studies forward. Front. Plant Sci. 2022, 13, 976627. [Google Scholar] [PubMed]
- Irgen-Gioro, S.; Yoshida, S.; Walling, V.; Chong, S. Fixation can change the appearance of phase separation in living cells. Elife 2022, 11, e79903. [Google Scholar] [CrossRef]
- Miné-Hattab, J.; Heltberg, M.; Villemeur, M.; Guedj, C.; Mora, T.; Walczak, A.M.; Dahan, M.; Taddei, A. Single molecule microscopy reveals key physical features of repair foci in living cells. Elife 2021, 10, e60577. [Google Scholar] [PubMed]
- Cui, Y.; Zhao, Y.; Lu, Y.; Su, X.; Chen, Y.; Shen, Y.; Lin, J.; Li, X. In vivo single particle tracking of the aquaporin AtPIP2;1 in stomata reveals cell type-specific dynamics. Plant Physiol. 2021, 185, 1666–1681. [Google Scholar] [CrossRef] [PubMed]
- Feng, Z.; Chen, X.; Wu, X.; Zhang, M. Formation of biological condensates via phase separation: Characteristics, analytical methods, and physiological implications. J. Biol. Chem. 2019, 294, 14823–14835. [Google Scholar] [CrossRef]
- Pandey, N.K.; Varkey, J.; Ajayan, A.; George, G.; Chen, J.; Langen, R. Fluorescent protein tagging promotes phase separation and alters the aggregation pathway of huntingtin exon-1. J. Biol. Chem. 2024, 300, 105585. [Google Scholar] [CrossRef]
- Schneider, R.; Blackledge, M.; Jensen, M.R. Elucidating binding mechanisms and dynamics of intrinsically disordered protein complexes using NMR spectroscopy. Curr. Opin. Struct. Biol. 2019, 54, 10–18. [Google Scholar]
- Kang, C. Applications of in-cell NMR in structural biology and drug discovery. Int. J. Mol. Sci. 2019, 20, 139. [Google Scholar] [CrossRef]
- Cedeño, C.; Pauwels, K.; Tompa, P. Protein delivery into plant cells: Toward in vivo structural biology. Front. Plant Sci. 2017, 8, 519. [Google Scholar] [CrossRef]
- Cuevas-Velazquez, C.L.; Vellosillo, T.; Guadalupe, K.; Schmidt, H.B.; Yu, F.; Moses, D.; Brophy, J.A.N.; Cosio-Acosta, D.; Das, A.; Wang, L.; et al. Intrinsically disordered protein biosensor tracks the physical-chemical effects of osmotic stress on cells. Nat. Commun. 2021, 12, 5438. [Google Scholar] [CrossRef]
- Kasianchuk, N.; Rzymski, P.; Kaczmarek, Ł. The biomedical potential of tardigrade proteins: A review. Biomed. Pharmacother. 2023, 158, 114063. [Google Scholar]
Figure 1.
Putative molecular shielding mechanisms for cryoprotective activity of intrinsically disordered proteins (IDPs). During freezing stress, the enzyme (i.e., lactate dehydrogenase) increases protein–protein contacts to cause aggregation, which inactivates enzyme activity. In the presence of IDPs (i.e., LEA proteins) during freezing stress, the IDP acts as a shield, interfering with the interaction of the enzyme and keeping it in an active state.
Figure 1.
Putative molecular shielding mechanisms for cryoprotective activity of intrinsically disordered proteins (IDPs). During freezing stress, the enzyme (i.e., lactate dehydrogenase) increases protein–protein contacts to cause aggregation, which inactivates enzyme activity. In the presence of IDPs (i.e., LEA proteins) during freezing stress, the IDP acts as a shield, interfering with the interaction of the enzyme and keeping it in an active state.

Figure 2.
Motif organization and induced folding of three representative plant IDPs, Lti30, StREM1.3, and ASR1. (A) Arabidopsis Lti30 is a dehydrin that contains six K segments flanked by histidine (H)-rich sequences. K segments form helical conformations upon dehydration or when positively charged lysine (K+) and histidine (H+) interact with the negatively charged head groups of membrane phospholipids. (B) Potato remorin StREM1.3 contains the N-terminal IDR domain and a coiled-coil (CC) region and the C-terminal anchor (CA). The lipid-induced folding of CA region interacts with lipid polar heads via positively charged lysine (K+). (C) The protein structure of tomato ASR1 contains a short N-terminal consensus sequence comprising histidine (H) residues and a conserved abscisic-acid–water deficit stress (ABA_WDS) domain based on https://www.ebi.ac.uk/interpro/protein/UniProt/Q08655/ (accessed on 28 October 2023). (D) Disordered motif organization of tomato ASR1 drawn according to the sequences published on Goldgur et al. [106]. The three disordered motifs are rich in histidine, lysine, and glutamic acid (HKE). ASR1 can gain ordered structures upon the binding of zinc (Zn) at the N-terminal motif. Zn binding at the central motif contributes to the DNA-binding activity of ASR1. H+, positively charged histidine; K+, positively charged lysine; IDR, intrinsically disordered region.
Figure 2.
Motif organization and induced folding of three representative plant IDPs, Lti30, StREM1.3, and ASR1. (A) Arabidopsis Lti30 is a dehydrin that contains six K segments flanked by histidine (H)-rich sequences. K segments form helical conformations upon dehydration or when positively charged lysine (K+) and histidine (H+) interact with the negatively charged head groups of membrane phospholipids. (B) Potato remorin StREM1.3 contains the N-terminal IDR domain and a coiled-coil (CC) region and the C-terminal anchor (CA). The lipid-induced folding of CA region interacts with lipid polar heads via positively charged lysine (K+). (C) The protein structure of tomato ASR1 contains a short N-terminal consensus sequence comprising histidine (H) residues and a conserved abscisic-acid–water deficit stress (ABA_WDS) domain based on https://www.ebi.ac.uk/interpro/protein/UniProt/Q08655/ (accessed on 28 October 2023). (D) Disordered motif organization of tomato ASR1 drawn according to the sequences published on Goldgur et al. [106]. The three disordered motifs are rich in histidine, lysine, and glutamic acid (HKE). ASR1 can gain ordered structures upon the binding of zinc (Zn) at the N-terminal motif. Zn binding at the central motif contributes to the DNA-binding activity of ASR1. H+, positively charged histidine; K+, positively charged lysine; IDR, intrinsically disordered region.

Figure 3.
Protein disorder is important in abiotic and biotic stress resistance in plants. (A) Intrinsically disordered proteins (IDPs) can undergo disorder-to-order transition and form an α-helix to bind cellular membranes, maintaining membrane stability under freezing stress. (B) IDPs can interact with membrane proteins such as transporters to mediate their functions under stress conditions. (C) During plant–pathogen interactions, IDPs mediate the molecular condensation of the cell wall–plasma membrane–cytoskeleton continuum for immunity signalling transduction. (D) IDPs and intrinsically disordered regions (IDRs) interact with the cytoskeletal actin filaments and microtubules for adaptive growth under water deficit and salinity stress. (E) IDPs can form a molecular shield to protect enzymes under stress conditions. (F) Metal ion binding of IDPs induces their conformational changes and facilitates their interaction with DNA. (G) Liquid–liquid phase separation (LLPS) of IDPs may regulate gene expression during plant immunity responses.
Figure 3.
Protein disorder is important in abiotic and biotic stress resistance in plants. (A) Intrinsically disordered proteins (IDPs) can undergo disorder-to-order transition and form an α-helix to bind cellular membranes, maintaining membrane stability under freezing stress. (B) IDPs can interact with membrane proteins such as transporters to mediate their functions under stress conditions. (C) During plant–pathogen interactions, IDPs mediate the molecular condensation of the cell wall–plasma membrane–cytoskeleton continuum for immunity signalling transduction. (D) IDPs and intrinsically disordered regions (IDRs) interact with the cytoskeletal actin filaments and microtubules for adaptive growth under water deficit and salinity stress. (E) IDPs can form a molecular shield to protect enzymes under stress conditions. (F) Metal ion binding of IDPs induces their conformational changes and facilitates their interaction with DNA. (G) Liquid–liquid phase separation (LLPS) of IDPs may regulate gene expression during plant immunity responses.

Table 2.
Studies of late embryogenesis abundant (LEA) proteins mentioned in Section 2.
Table 2.
Studies of late embryogenesis abundant (LEA) proteins mentioned in Section 2.
| Plant Species | Finding | Topics | References |
|---|---|---|---|
| Rice (Oryza sativa) | Identification of 34 rice LEA genes and their transcript analysis under untreated, abscisic acid, osmotic, and salinity stress conditions. | Genomics | [38] |
| Tomato (Solanum lycopersicum) | Identification of 60 tomato LEA genes and their transcription and expression patterns in various tissues and under abiotic stress and phytohormone treatments. Duplication event in evolution of the LEA family. | Genomics | [39,41] |
| Characteristics of the LEA proteins including disorder tendency and localization. | Proteomics | [41] | |
| Arabidopsis (Arabidopsis thaliana) | Identification of 51 Arabidopsis LEA genes and their transcription patterns at different developmental stages, in different plant organs, and under different stress and hormone treatments. Respective promoter elements induced by abiotic stress. | Genomics | [40] |
| Classification of Arabidopsis LEA proteins and their subcellular localization. | Cell biology | [60] | |
| 60 plant species ranging from green algae to angiosperms | Comprehensive synteny and phylogenetic analyses of LEA genes across 60 complete plant genomes showed their evolution and diversification. | Genomics | [42] |
| Orchid (Dendrobium officinale) | Identification of Dendrobium officinale LEA genes and their expression patterns under abiotic stress treatments. Classification of DfLEA proteins and their predicted subcellular localization. | Genomics | [61] |
| Xerophyta schlechteri | Enzyme protection function of XsLEA proteins under stressful conditions. Expression of XsLEAs increased abiotic stress tolerance in E. coli and Arabidopsis. | Stress physiology | [57] |
| Alfalfa (Medicago sativa) | Expression of Medicago sativa MsLEA-D34 increased plant tolerance to osmotic and salt stresses and caused Arabidopsis early flowering under drought and well-watered conditions. | Stress physiology | [58] |
| Bermudagrass (Cynodon dactylon) | Expression of Cynodon dactylon CdDHN4-L and CdDHN4-S increased salt tolerance in Arabidopsis, enzyme protection function, and disordered character. | Stress physiology | [59] |
| Common wheat (Triticum aestivum), durum wheat (T. durum), barley (Hordeum vulgare) | A comparison of dehydrins in common wheat, durum wheat and barley in the context of expression patterns at transcript and protein levels and their possible functions when exposed to various abiotic stress factors. | Stress physiology | [15] |
| Various | A summary of structural characterization of plant LEA proteins and their binding modes. | Structural biology | [22] |
| Various | A summary of plant LEA proteins from characterization to their functions in stress responses. | Stress physiology | [35] |
| Various | A summary of LEA functions during seed maturation and seed desiccation tolerance. | Stress physiology | [36] |
| Arabidopsis (Arabidopsis thaliana) | A discussion of LEA functions in stabilizing membranes or sensitive enzymes during dehydration. | Stress physiology | [37] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Hsiao, A.-S. Protein Disorder in Plant Stress Adaptation: From Late Embryogenesis Abundant to Other Intrinsically Disordered Proteins. Int. J. Mol. Sci. 2024, 25, 1178. https://doi.org/10.3390/ijms25021178
AMA Style
Hsiao A-S. Protein Disorder in Plant Stress Adaptation: From Late Embryogenesis Abundant to Other Intrinsically Disordered Proteins. International Journal of Molecular Sciences. 2024; 25(2):1178. https://doi.org/10.3390/ijms25021178
Chicago/Turabian StyleHsiao, An-Shan. 2024. "Protein Disorder in Plant Stress Adaptation: From Late Embryogenesis Abundant to Other Intrinsically Disordered Proteins" International Journal of Molecular Sciences 25, no. 2: 1178. https://doi.org/10.3390/ijms25021178
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.