Salivary Histatin 1 and 2 Are Targeted to Mitochondria and Endoplasmic Reticulum in Human Cells


Abstract
:1. Introduction
2. Materials and Methods
2.1. Solid-Phase Peptide Synthesis
2.2. Cell Culture
2.3. Fluorescence Microscopy
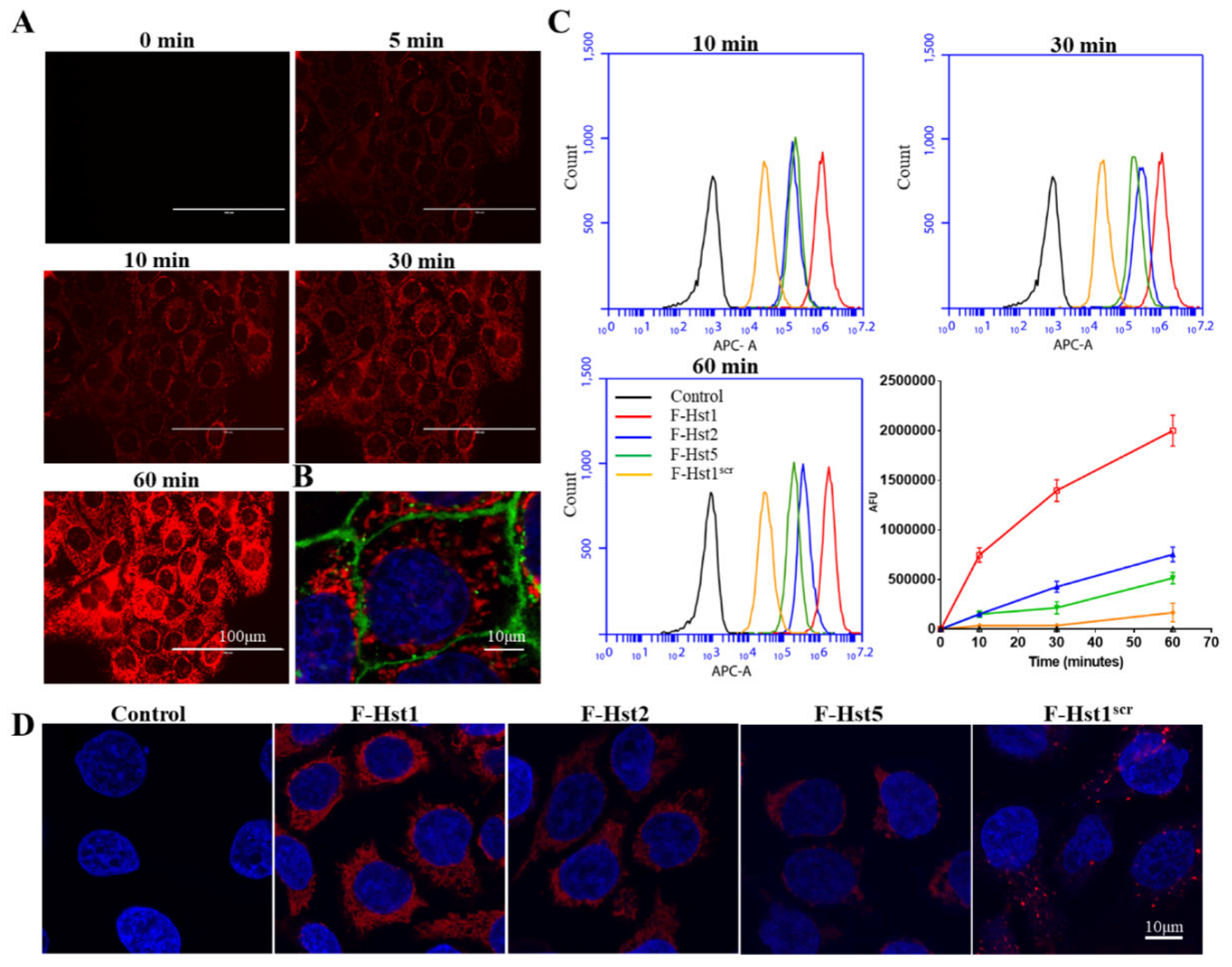
2.4. Flow Cytometry
2.5. Cellular Internalization of the F-Hsts
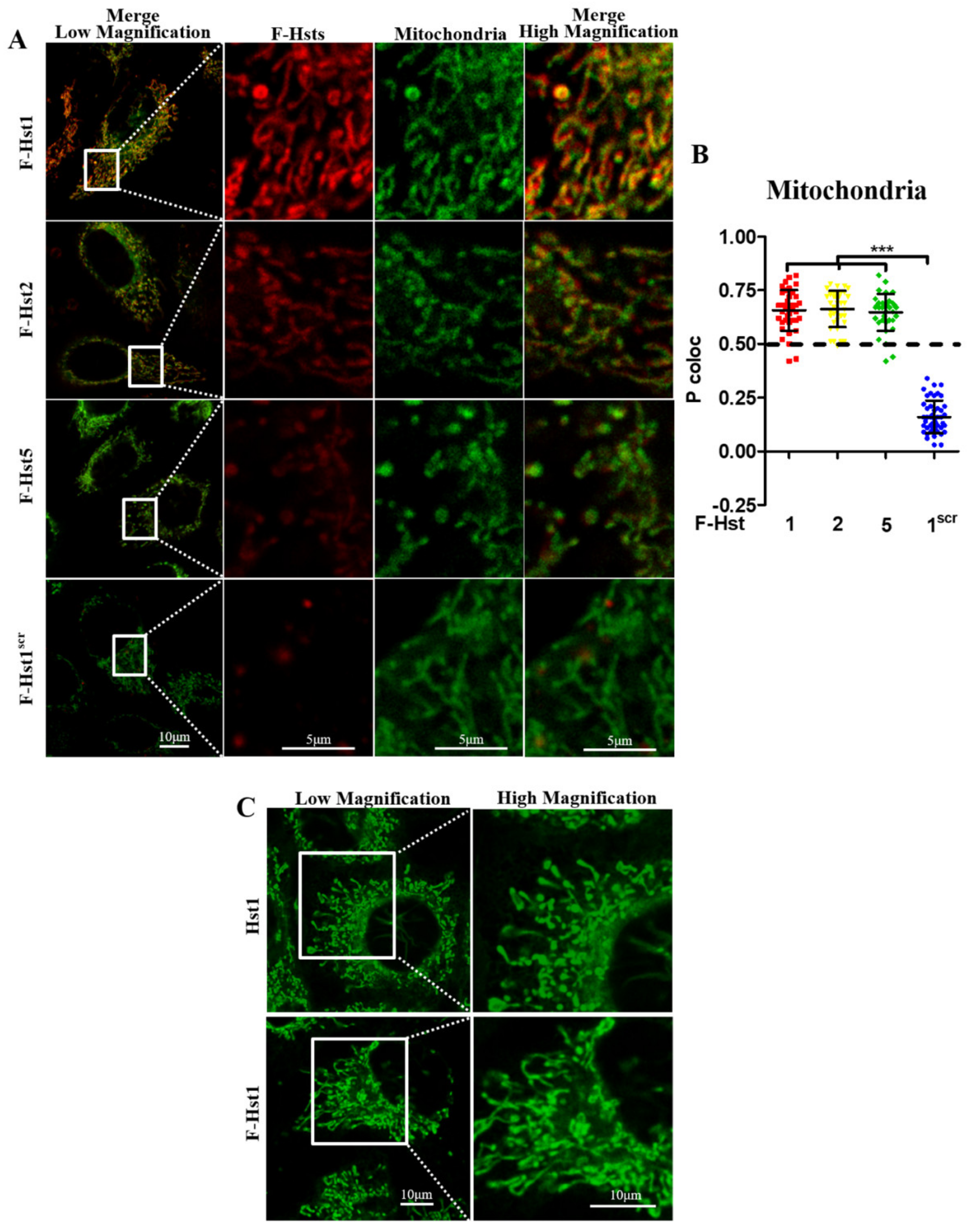
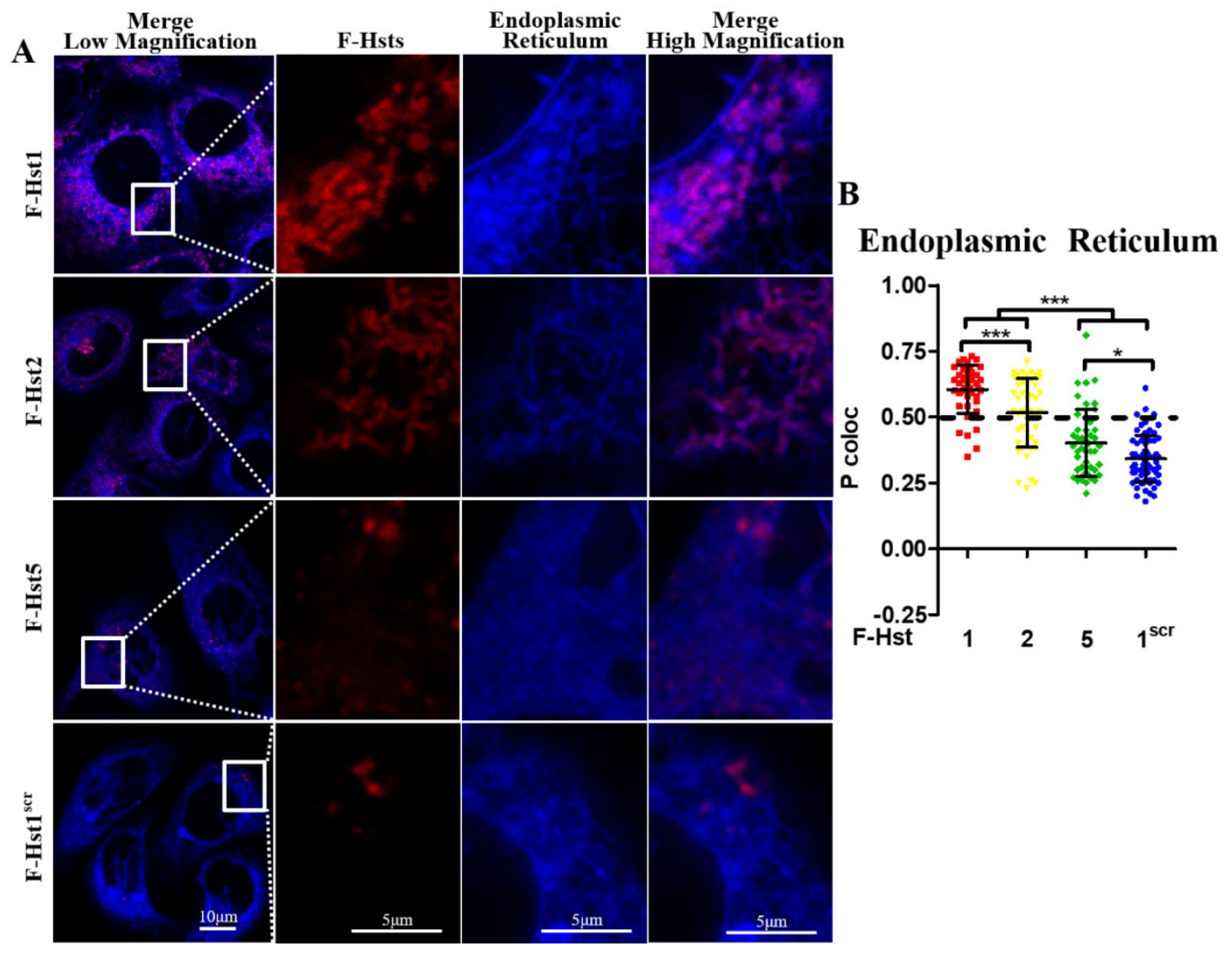
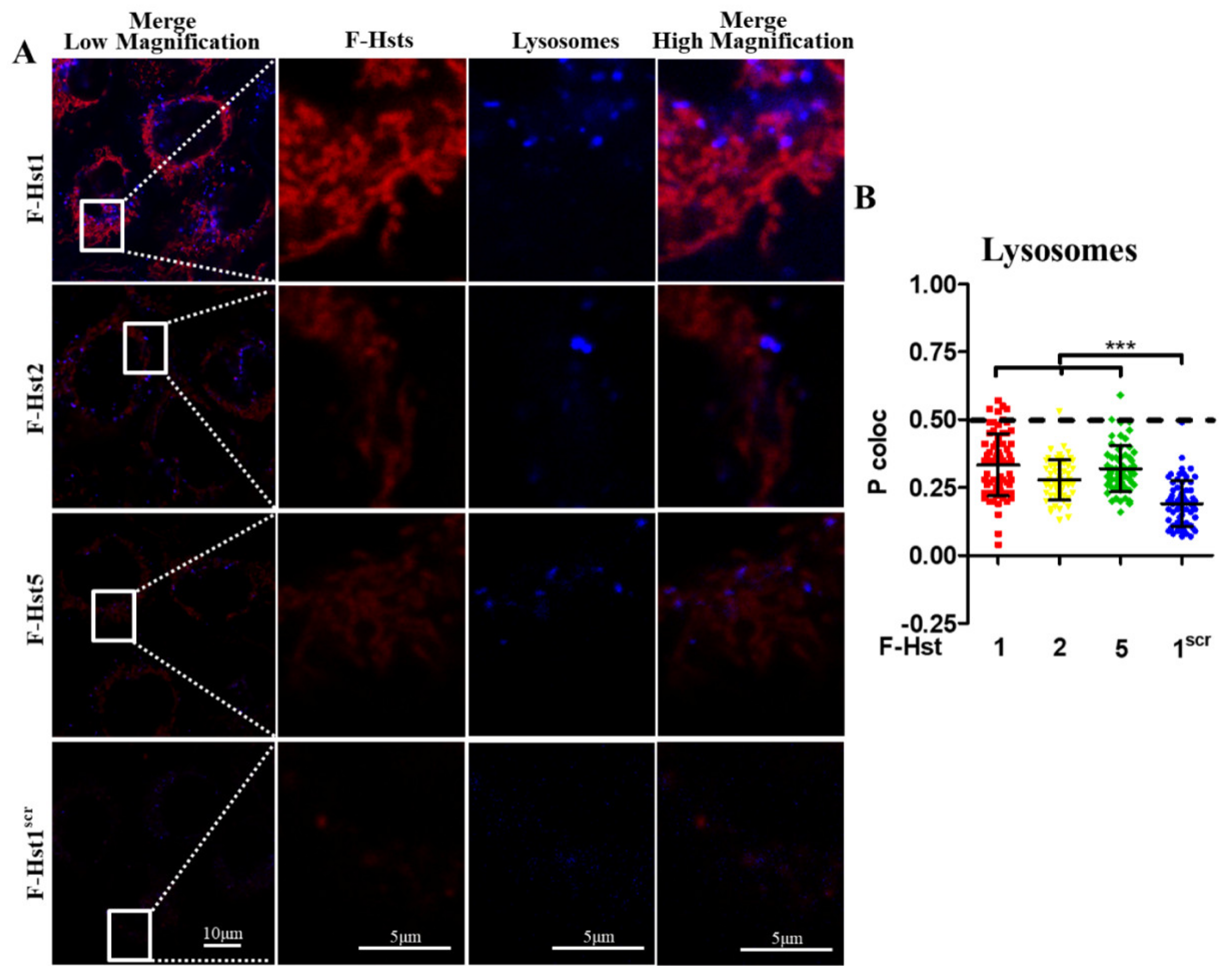
2.6. Subcellular Localization of the F-Hsts
2.7. Cellular Metabolic Activity Assay
2.8. Statistical Analysis
3. Results
3.1. Uptake Dynamics of the F-Hsts
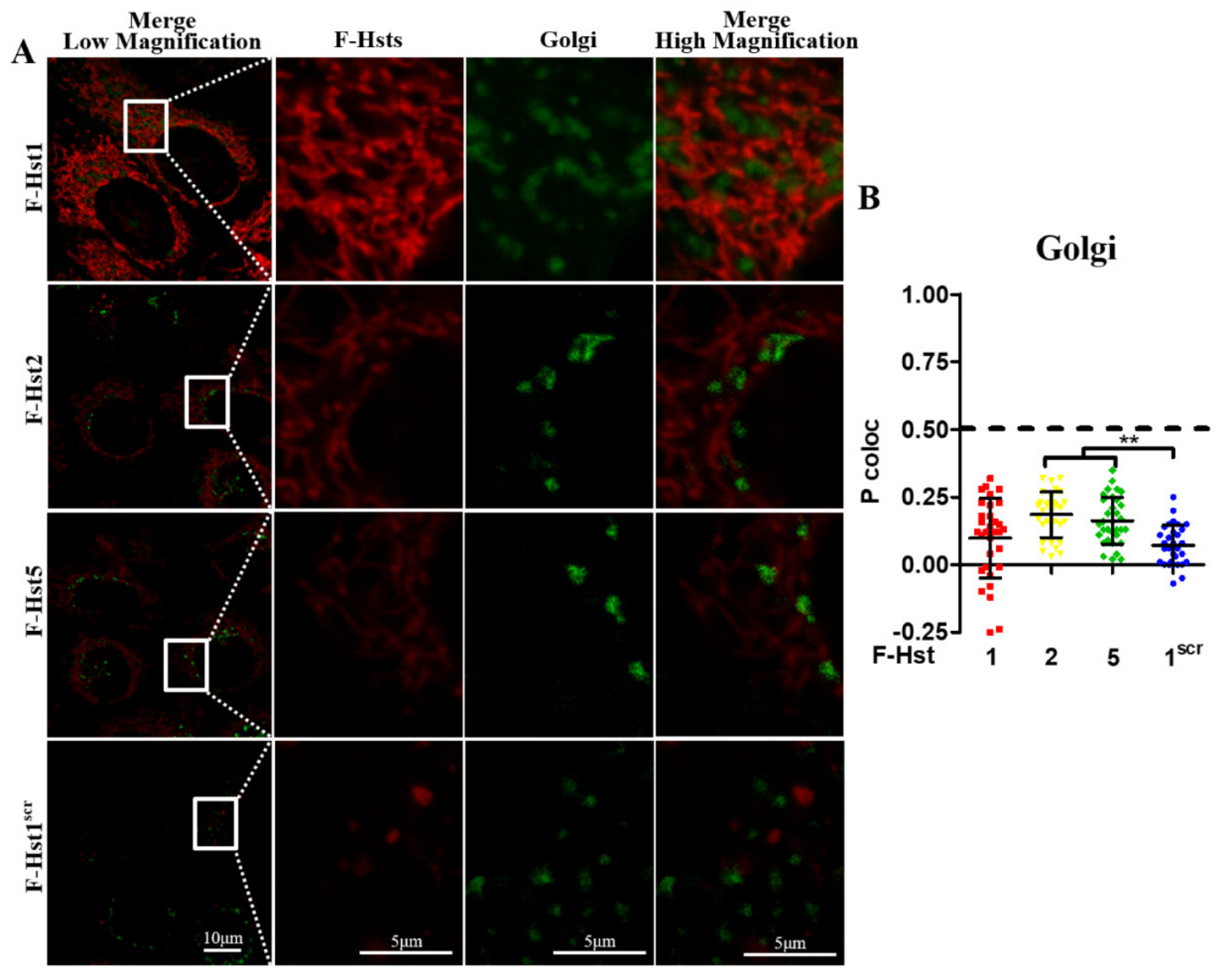
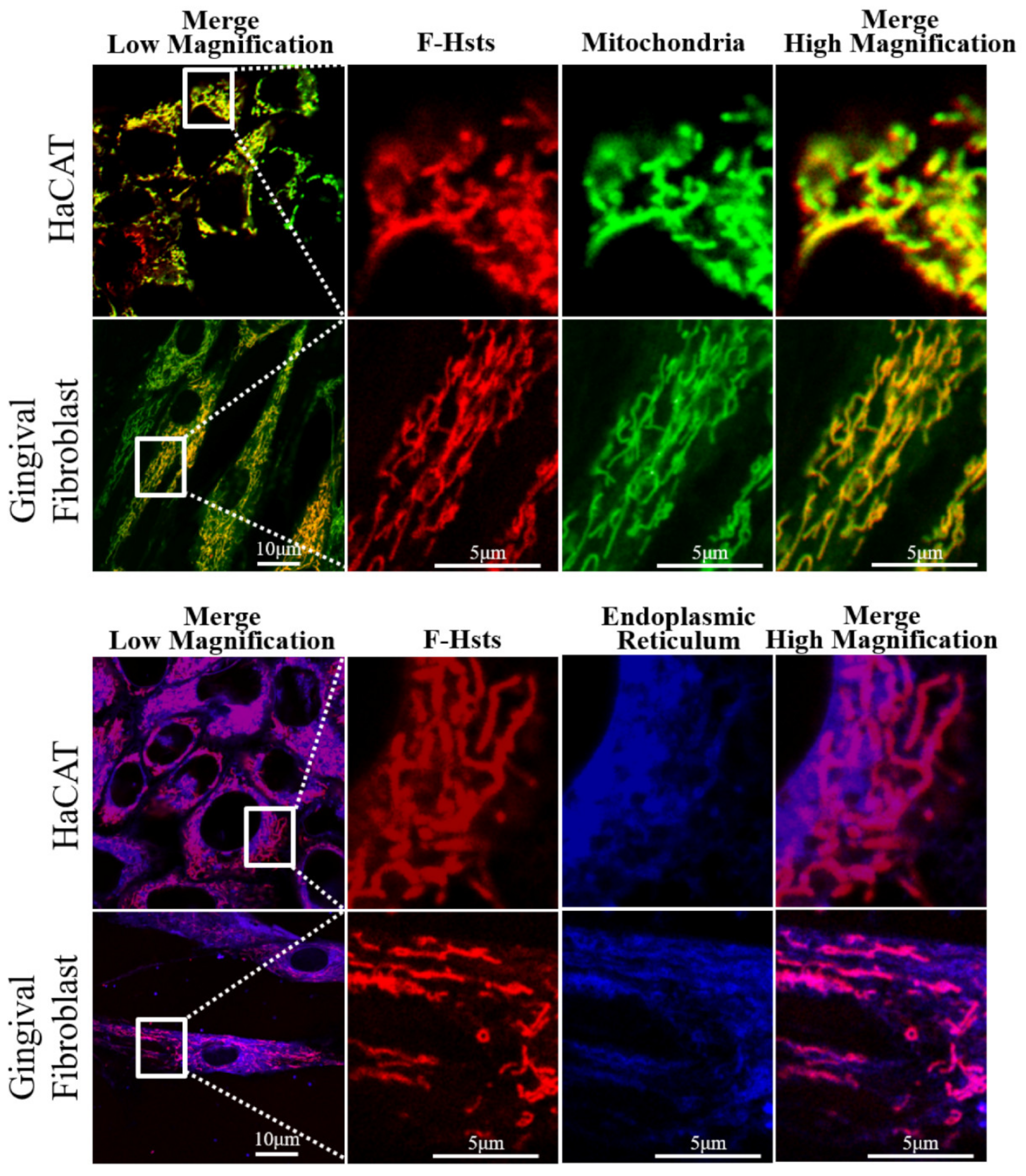
3.2. Subcellular Localization of the F-Hsts
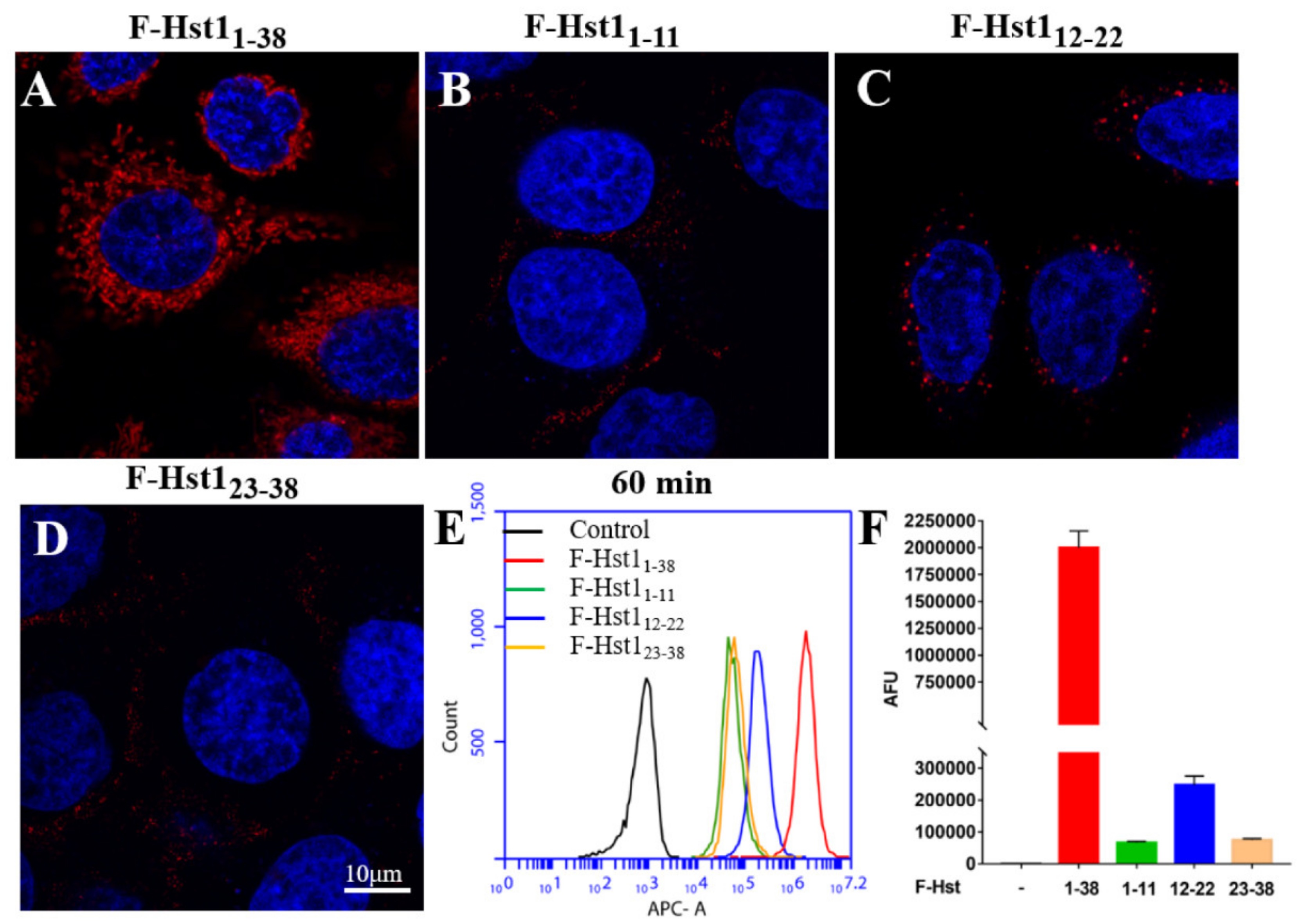
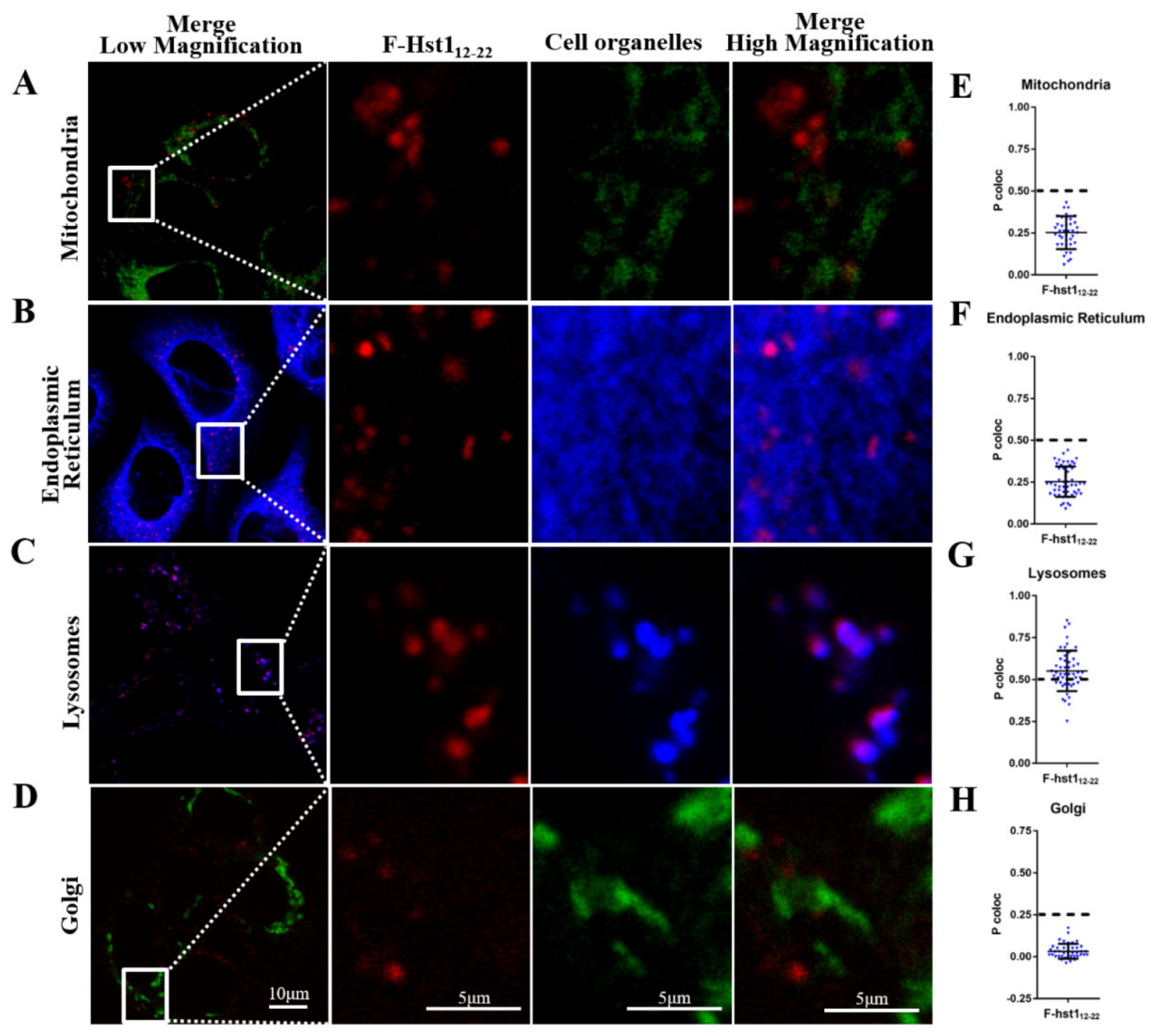
3.3. Uptake Dynamics and Subcellular Target of Truncated F-Hsts, F-Hst11-11, F-Hst112-22 and F-Hst123-38
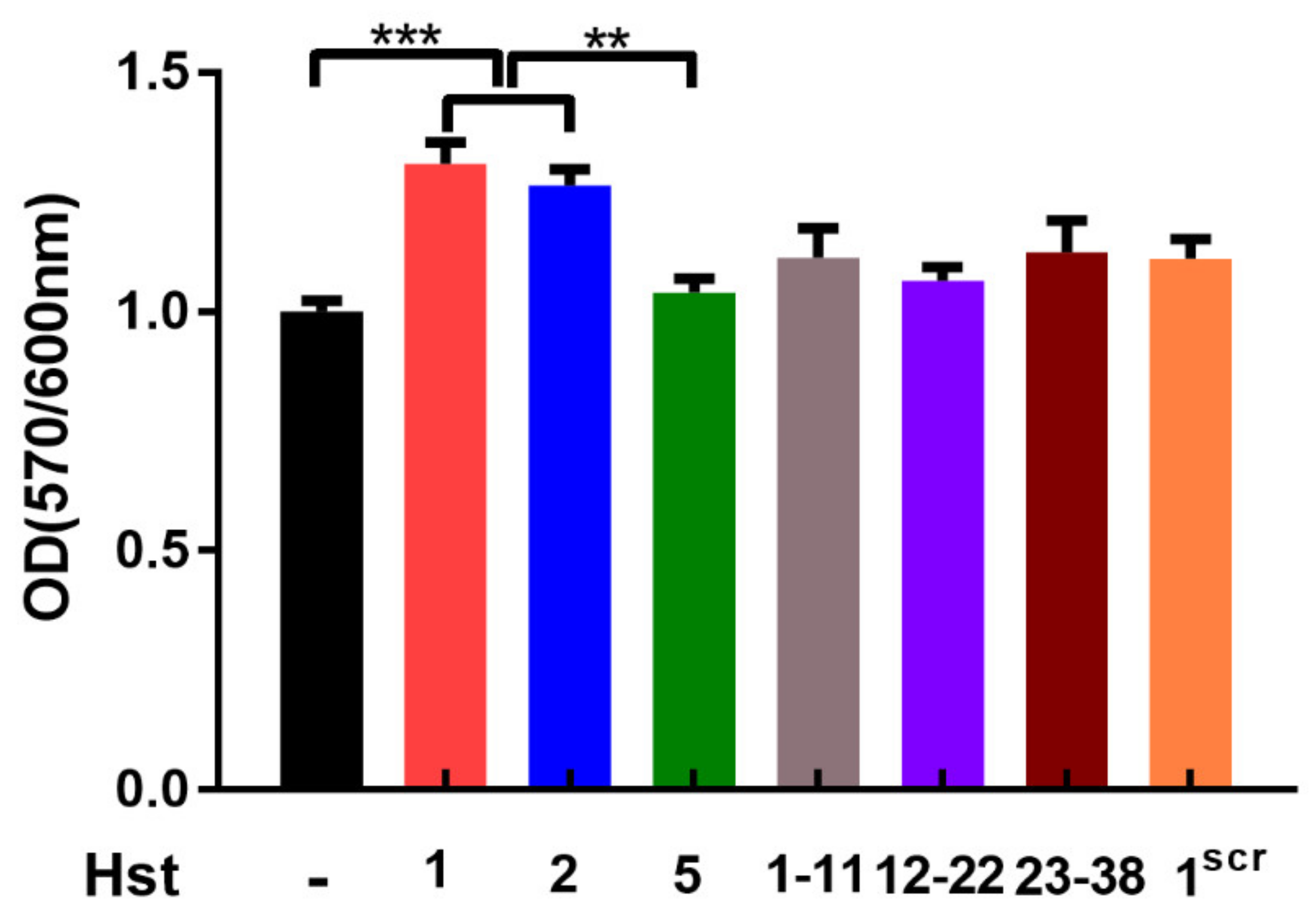
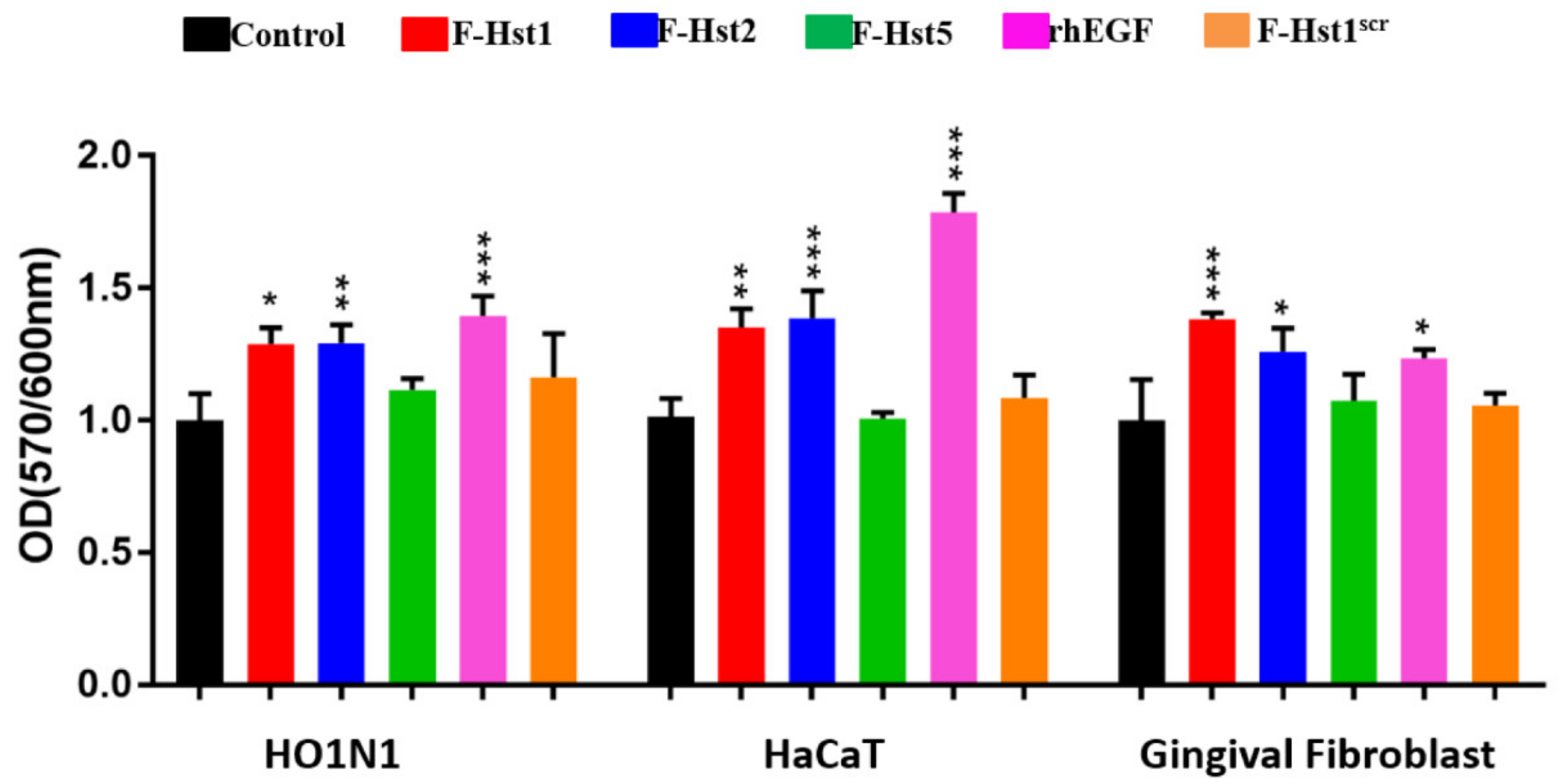
3.4. Cellular Metabolic Activity
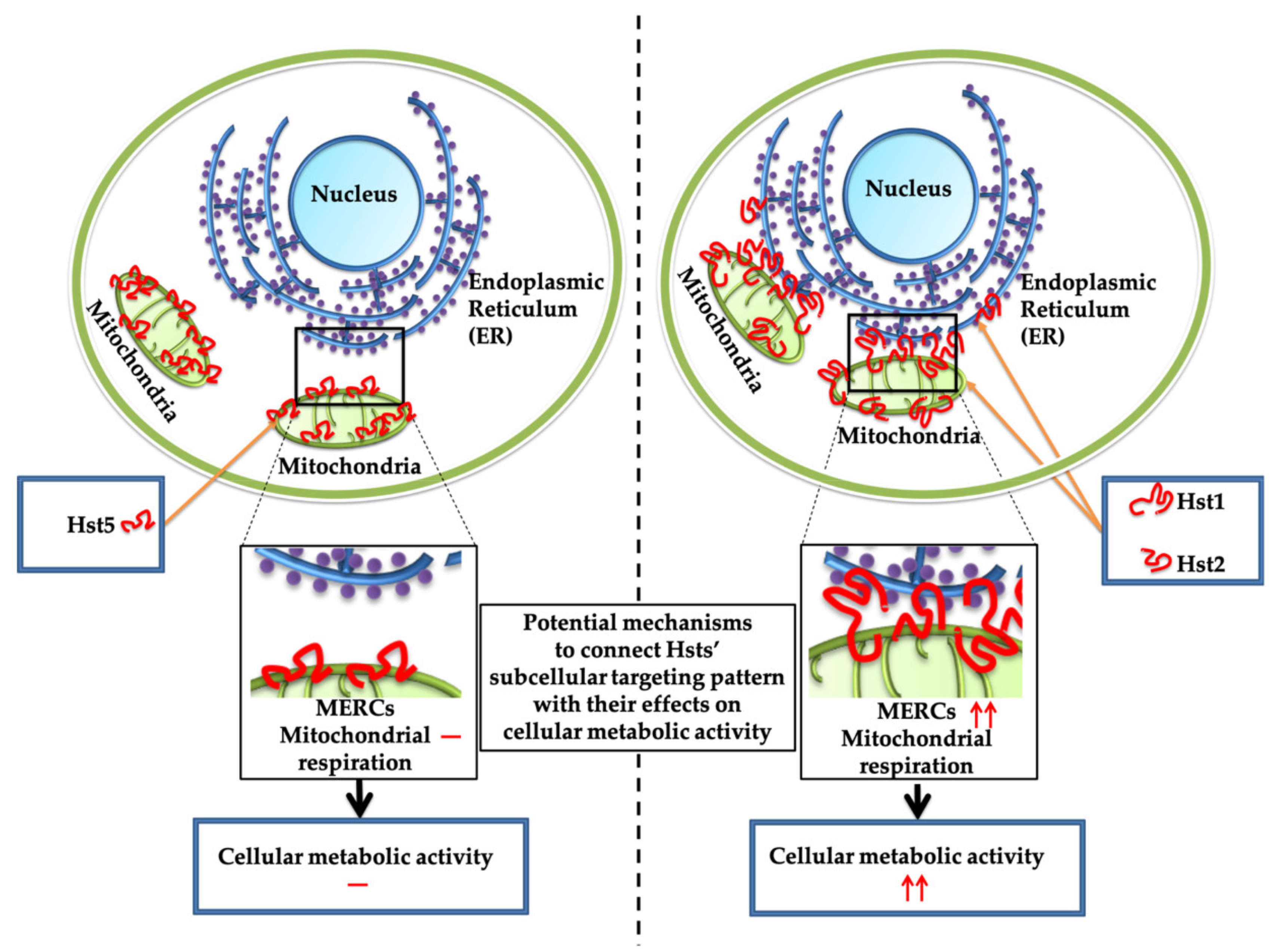
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Vander Spek, J.C.; Offner, G.D.; Troxler, R.F.; Oppenheim, F.G. Molecular cloning of human submandibular histatins. Arch. Oral. Biol. 1990, 35, 137–143. [Google Scholar] [CrossRef]
- Oudhoff, M.J.; Bolscher, J.G.; Nazmi, K.; Kalay, H.; van ‘t Hof, W.; Amerongen, A.V.; Veerman, E.C. Histatins are the major wound-closure stimulating factors in human saliva as identified in a cell culture assay. Faseb. J. 2008, 22, 3805–3812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oudhoff, M.J.; Kroeze, K.L.; Nazmi, K.; van den Keijbus, P.A.; van ‘t Hof, W.; Fernandez-Borja, M.; Hordijk, P.L.; Gibbs, S.; Bolscher, J.G.; Veerman, E.C. Structure-activity analysis of histatin, a potent wound healing peptide from human saliva: Cyclization of histatin potentiates molar activity 1000-fold. Faseb. J. 2009, 23, 3928–3935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oudhoff, M.J.; van den Keijbus, P.A.; Kroeze, K.L.; Nazmi, K.; Gibbs, S.; Bolscher, J.G.; Veerman, E.C. Histatins enhance wound closure with oral and non-oral cells. J. Dent. Res. 2009, 88, 846–850. [Google Scholar] [CrossRef] [PubMed]
- Van Dijk, I.A.; Beker, A.F.; Jellema, W.; Nazmi, K.; Wu, G.; Wismeijer, D.; Krawczyk, P.M.; Bolscher, J.G.; Veerman, E.C.; Stap, J. Histatin 1 Enhances Cell Adhesion to Titanium in an Implant Integration Model. J. Dent. Res. 2017, 96, 430–436. [Google Scholar] [CrossRef] [PubMed]
- Van Dijk, I.A.; Ferrando, M.L.; van der Wijk, A.E.; Hoebe, R.A.; Nazmi, K.; de Jonge, W.J.; Krawczyk, P.M.; Bolscher, J.G.M.; Veerman, E.C.I.; Stap, J. Human salivary peptide histatin-1 stimulates epithelial and endothelial cell adhesion and barrier function. Faseb. J. 2017, 31, 3922–3933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Dijk, I.A.; Nazmi, K.; Bolscher, J.G.; Veerman, E.C.; Stap, J. Histatin-1, a histidine-rich peptide in human saliva, promotes cell-substrate and cell-cell adhesion. Faseb. J. 2015, 29, 3124–3132. [Google Scholar] [CrossRef]
- Veerman, E.C.; Valentijn-Benz, M.; Nazmi, K.; Ruissen, A.L.; Walgreen-Weterings, E.; van Marle, J.; Doust, A.B.; van’t Hof, W.; Bolscher, J.G.; Amerongen, A.V. Energy depletion protects Candida albicans against antimicrobial peptides by rigidifying its cell membrane. J. Biol. Chem. 2007, 282, 18831–18841. [Google Scholar] [CrossRef] [Green Version]
- Veerman, E.C.I.; Nazmi, K.; van Hof, W.; Bolscher, J.G.M.; den Hertog, A.L.; Amerongen, A.V.N. Reactive oxygen species play no role in the candidacidal activity of the salivary antimicrobial peptide histatin 5. Biochem. J. 2004, 381, 447–452. [Google Scholar] [CrossRef] [Green Version]
- Helmerhorst, E.J.; Breeuwer, P.; van’t Hof, W.; Walgreen-Weterings, E.; Oomen, L.C.; Veerman, E.C.; Amerongen, A.V.; Abee, T. The cellular target of histatin 5 on Candida albicans is the energized mitochondrion. J. Biol. Chem. 1999, 274, 7286–7291. [Google Scholar] [CrossRef] [Green Version]
- Den Hertog, A.L.; Wong Fong Sang, H.W.; Kraayenhof, R.; Bolscher, J.G.; Van’t Hof, W.; Veerman, E.C.; Nieuw Amerongen, A.V. Interactions of histatin 5 and histatin 5-derived peptides with liposome membranes: Surface effects, translocation and permeabilization. Biochem. J. 2004, 379, 665–672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torres, P.; Diaz, J.; Arce, M.; Silva, P.; Mendoza, P.; Lois, P.; Molina-Berrios, A.; Owen, G.I.; Palma, V.; Torres, V.A. The salivary peptide histatin-1 promotes endothelial cell adhesion, migration, and angiogenesis. Faseb. J. 2017, 31, 4946–4958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shah, D.; Ali, M.; Shukla, D.; Jain, S.; Aakalu, V.K. Effects of histatin-1 peptide on human corneal epithelial cells. PLoS ONE 2017, 12, e0178030. [Google Scholar] [CrossRef] [PubMed]
- Castro, M.; Torres, P.; Solano, L.; Cordova, L.A.; Torres, V.A. Histatin-1 counteracts the cytotoxic and antimigratory effects of zoledronic acid in endothelial and osteoblast-like cells. J. Periodontol. 2019, 90, 766–774. [Google Scholar] [CrossRef]
- Huang, G.Q.; Yi, G.G.; Wu, L.W.; Feng, S.F.; Wu, W.; Peng, L.; Yi, R.W.; Ma, W.; Lu, X. Protective effect of histatin 1 against ultraviolet-induced damage to human corneal epithelial cells. Exp. Ther. Med. 2018, 15, 679–684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oppenheim, F.G.; Xu, T.; McMillian, F.M.; Levitz, S.M.; Diamond, R.D.; Offner, G.D.; Troxler, R.F. Histatins, a novel family of histidine-rich proteins in human parotid secretion. Isolation, characterization, primary structure, and fungistatic effects on Candida albicans. J. Biol. Chem. 1988, 263, 7472–7477. [Google Scholar]
- Xu, T.; Levitz, S.M.; Diamond, R.D.; Oppenheim, F.G. Anticandidal activity of major human salivary histatins. Infect. Immun. 1991, 59, 2549–2554. [Google Scholar] [CrossRef] [Green Version]
- Helmerhorst, E.J.; Reijnders, I.M.; van’t Hof, W.; Simoons-Smit, I.; Veerman, E.C.; Amerongen, A.V. Amphotericin B- and fluconazole-resistant Candida spp., Aspergillus fumigatus, and other newly emerging pathogenic fungi are susceptible to basic antifungal peptides. Antimicrob. Agents Chemother. 1999, 43, 702–704. [Google Scholar] [CrossRef] [Green Version]
- Lin, A.L.; Shi, Q.; Johnson, D.A.; Patterson, T.F.; Rinaldi, M.G.; Yeh, C.K. Further characterization of human salivary anticandidal activities in a human immunodeficiency virus-positive cohort by use of microassays. Clin. Diagn. Lab. Immunol. 1999, 6, 851–855. [Google Scholar] [CrossRef] [Green Version]
- Mochon, A.B.; Liu, H. The antimicrobial peptide histatin-5 causes a spatially restricted disruption on the Candida albicans surface, allowing rapid entry of the peptide into the cytoplasm. PLoS Pathog. 2008, 4, e1000190. [Google Scholar] [CrossRef] [Green Version]
- Bolscher, J.G.; Oudhoff, M.J.; Nazmi, K.; Antos, J.M.; Guimaraes, C.P.; Spooner, E.; Haney, E.F.; Garcia Vallejo, J.J.; Vogel, H.J.; van’t Hof, W.; et al. Sortase A as a tool for high-yield histatin cyclization. Faseb. J. 2011, 25, 2650–2658. [Google Scholar] [CrossRef] [PubMed]
- Gyurko, C.; Lendenmann, U.; Troxler, R.F.; Oppenheim, F.G. Candida albicans mutants deficient in respiration are resistant to the small cationic salivary antimicrobial peptide histatin 5. Antimicrob. Agents Chemother. 2000, 44, 348–354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gyurko, C.; Lendenmann, U.; Helmerhorst, E.J.; Troxler, R.F.; Oppenheim, F.G. Killing of Candida albicans by histatin 5: Cellular uptake and energy requirement. Antonie Van Leeuwenhoek 2001, 79, 297–309. [Google Scholar] [CrossRef] [PubMed]
- Luque-Ortega, J.R.; van’t Hof, W.; Veerman, E.C.; Saugar, J.M.; Rivas, L. Human antimicrobial peptide histatin 5 is a cell-penetrating peptide targeting mitochondrial ATP synthesis in Leishmania. Faseb. J. 2008, 22, 1817–1828. [Google Scholar] [CrossRef] [Green Version]
- Oudhoff, M.J. Discovery of the Wound-Healing Capacity of Salivary Histatins. Ph.D. Thesis, Vrije Universiteit, Amsterdam, The Netherlands, 7 December 2010. [Google Scholar]
- Szymanski, J.; Janikiewicz, J.; Michalska, B.; Patalas-Krawczyk, P.; Perrone, M.; Ziolkowski, W.; Duszynski, J.; Pinton, P.; Dobrzyn, A.; Wieckowski, M.R. Interaction of Mitochondria with the Endoplasmic Reticulum and Plasma Membrane in Calcium Homeostasis, Lipid Trafficking and Mitochondrial Structure. Int. J. Mol. Sci. 2017, 18, 1576. [Google Scholar] [CrossRef]
- Da Silva, A.F.; Mariotti, F.R.; Maximo, V.; Campello, S. Mitochondria dynamism: Of shape, transport and cell migration. Cell Mol. Life Sci. 2014, 71, 2313–2324. [Google Scholar] [CrossRef]
- Paupe, V.; Prudent, J. New insights into the role of mitochondrial calcium homeostasis in cell migration. Biochem. Biophys. Res. Commun. 2018, 500, 75–86. [Google Scholar] [CrossRef]
- Alshaabi, H.; Heininger, M.; Cunniff, B. Dynamic regulation of subcellular mitochondrial position for localized metabolite levels. J. Biochem. 2019. [Google Scholar] [CrossRef]
- Bravo, R.; Vicencio, J.M.; Parra, V.; Troncoso, R.; Munoz, J.P.; Bui, M.; Quiroga, C.; Rodriguez, A.E.; Verdejo, H.E.; Ferreira, J.; et al. Increased ER-mitochondrial coupling promotes mitochondrial respiration and bioenergetics during early phases of ER stress. J. Cell Sci. 2011, 124, 2143–2152. [Google Scholar] [CrossRef] [Green Version]
- Tokuo, H.; Mabuchi, K.; Ikebe, M. The motor activity of myosin-X promotes actin fiber convergence at the cell periphery to initiate filopodia formation. J. Cell Biol. 2007, 179, 229–238. [Google Scholar] [CrossRef]
- Romero, S.; Didry, D.; Larquet, E.; Boisset, N.; Pantaloni, D.; Carlier, M.F. How ATP hydrolysis controls filament assembly from profilin-actin: Implication for formin processivity. J. Biol. Chem. 2007, 282, 8435–8445. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptides | Amino Acid Sequences of Hst Variants |
|---|---|
| (F-) Hst11-38 | DSHEKRHHGYRRKFHEKHHSHREFPFYGDYGSNYLYDN |
| (F-) Hst2 | RKFHEKHHSHREFPFYGDYGSNYLYDN |
| (F-) Hst5 | DSHAKRHHGYKRKFHEKHHSHRGY |
| (F-) Hst11-11 | DSHEKRHHGYR |
| (F-) Hst112-22 | RKFHEKHHSHR |
| (F-) Hst123-38 | EFPFYGDYGSNYLYDN |
| (F-) Hst1scr | SDHSRHEEFKPRFHYHGGDYYRGRSKNFYHLEYKDHNH |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, D.; Sun, W.; Nazmi, K.; Veerman, E.C.I.; Bikker, F.J.; Jaspers, R.T.; Bolscher, J.G.M.; Wu, G. Salivary Histatin 1 and 2 Are Targeted to Mitochondria and Endoplasmic Reticulum in Human Cells. Cells 2020, 9, 795. https://doi.org/10.3390/cells9040795
Ma D, Sun W, Nazmi K, Veerman ECI, Bikker FJ, Jaspers RT, Bolscher JGM, Wu G. Salivary Histatin 1 and 2 Are Targeted to Mitochondria and Endoplasmic Reticulum in Human Cells. Cells. 2020; 9(4):795. https://doi.org/10.3390/cells9040795
Chicago/Turabian StyleMa, Dandan, Wei Sun, Kamran Nazmi, Enno C. I. Veerman, Floris J. Bikker, Richard T. Jaspers, Jan G. M. Bolscher, and Gang Wu. 2020. "Salivary Histatin 1 and 2 Are Targeted to Mitochondria and Endoplasmic Reticulum in Human Cells" Cells 9, no. 4: 795. https://doi.org/10.3390/cells9040795